
Ovary
The ovary is a most peculiar organ, quite unlike any other. Hopefully, you have by now become familiar with some of the general patterns of tissue organization (e.g., hollow mucosal organs such as trachea and intestine; glandular organs such as kidney, liver and pancreas; lymphoid organs such as spleen, lymph nodes and thymus, etc). But the ovary is in a category of its own.
Even the concept of four basic tissue types must be stretched to fit ovarian tissue -- so we won't even try to assign tissue types to ovarian stromal and granulosa cells.
The outside (peritoneal) surface of the ovary is covered not by a standard serosa with simple squamous mesothelium but rather by a peculiar simple cuboidal-to-columnar mesothelium (focally pseudostratified). This mesothelium is sometimes unhelpfully called "germinal epithelium" although it does not contain germ cells.
The bulk of the ovary, both cortex and medulla, consists of stroma. Interspersed in the stroma are follicles, which contain oocytes.
Ovarian stroma consists of peculiar spindle-shaped stromal cells similar to fibroblasts and arranged into a characteristic whorled texture. Unlike ordinary connective tissue, ovarian stroma is highly cellular while supporting fibers (both reticular fibers and ordinary collagen) are inconspicuous. Stromal cells associated with maturing follicles may acquire endocrine function and secrete estrogens. The entire ovarian stroma is highly vascular.
The outer portion of the ovary is called the cortex. The inner portion is called the medulla. There is some ill-defined difference in stromal texture between cortex and medulla, but the main distinction is that the cortex includes all of the ovarian follicles while the medulla contains the larger blood vessels.
Embedded in the stroma of the ovarian cortex are many follicles (numbering several hundred thousand). Each follicle contains a single oocyte surrounded by granulosa cells. Granulosa cells are separated from the surrounding stroma by a basement membrane.
All of the follicles (and oocytes) that a woman will ever have are present in the ovary at birth (at least, according to conventional accounts; recent research [Nature Medicine 2012] has called this into question).
At any given time, most of the follicles remain in a resting condition. These are called primordial follicles.
During each monthly cycle, a few follicles are activated by follicle stimulating hormone (FSH) from the pituitary and begin progressing toward ovulation, through stages called primary follicles, secondary follicles, and tertiary or Graafian follicles.
(Since these "stages" represent arbitrary points in a continuum, there is some flexibility in identifying the intermediates.) Typically only one follicle completes the process in each cycle. Other follicles which had begun the process of maturation then degenerate, appearing temporarily as atretic follicles.
Most easily recognized are primordial follicles, which have a single layer of thin (squamous) granulosa cells around the oocyte, and Graafian follicles , which include a fluid-filled space or antrum. Primary follicles have one or more layers of cuboidal granulosa cells. Secondary follicles are larger (with more granulosa cells) and with surrounding stromal tissue organized into internal and external thecal layers. Note that any mature (Graafian) follicle is so large that an arbitrary section is unlikely to include the oocyte, or even the cumulus oophorus, even though such features are always shown in textbook pictures.
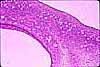
During follicle maturation, the granulosa cells proliferate to form many layers around the oocyte. A boundary layer, the zona pellucida, appears between the oocyte and the granulosa cells. Stromal cells surrounding the follicle differentiate into the theca interna and the theca externa. Cells of the theca interna become cuboidal steroid-producing cells (containing lipid droplets, extensive smooth endoplasmic reticulum, and mitochondria having tubular cristae). Androgen from thecal cells is processed to estrogen in granulosa cells; the estrogen then enters circulation via blood vessels which form in the theca. Cells of the theca externa have no known endocrine function.
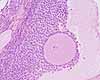
Eventually the layers of granulosa cells split apart to form a fluid-filled space called the antrum. At this stage it is called a tertiary or Graafian follicle (named after Reinier de Graaf, b. 1641). The mass of granulosa cells which continue to surround the follicle is called the "egg-bearing hill" or cumulus oophorus.
At ovulation, the follicle ruptures to release the oocyte, (with its associated "crown" of attached granulosa cells, the corona radiata.
Following ovulation and under the influence of luteinizing hormone, the ruptured follicle reorganizes into a
specialized endocrine structure called the corpus luteum ("yellow body"). The cuboidal granulosa cells enlarge and form ill-defined cords around vascular spaces (a process representing rapid growth of a temporary endocrine organ). These granulosa lutein cells assume a characteristic steroid-secreting appearance, with lipid droplets, extensive smooth endoplasmic reticulum, and mitochondria having tubular cristae. Thecal lutein cells are less obvious, smaller cuboidal cells associated with the fibrous septa of the corpus. The granulosa lutein cells secrete progesterone to maintain the uterus in a receptive phase and, if implantation occurs, continue to secrete progesterone through pregnancy.
The rapid development of a corpus luteum is quite curious; Here is a more detailed description, adapted from Weiss' Histology, 5th ed., pp. 923-6:
"... The transformation of the ovulatory follicle into a corpus luteum involves, first, the depolymerization of the basement membrane that originally separated the granulosa cell layers from the theca. This depolymerization allows the connective tissue cells and blood vessels to invade the srtatum granulosum... The granulosa cells that will become the luteal cells ... begin to undergo cytomorphosis even before ovulation [under the influence of LH]. ...Mitoses are seldom seen in the glandular parenchymal cells ... but are noted in the rapidly developing endothelium of the blood vessels which invade it.
"The luteal cells enlarge, become polyhedral, and are filled with lipid droplets... In ordinary histological preparations the cytoplasm of the luteal cells contains numerous empty vacuoles whose lipid contents are dissolved by the organic solvents used in processing the tissues. It is not known whether the several types of luteal cells that have been described are transitional stages in the differentiation of a single cell type or represent distinct cell species. An interesting feature of the developing corpus luteum is the rapid invasion of the stratum granulosum with connective tissue elements of the theca interna and sprouts of capillary endothelium. The connective tissue forms a delicate reticulum supporting the luteal cells. A complete rete network of capillaries forms rapidly throughout the gland. With time, larger blood vessels are formed. The formation of the vascular network in the developing corpus luteum is remarkably similar to that seen during the development of the vascular supply in any embryonic organ.
"In electron micrographs of the corpus luteum, the luteal cells have mitochondria with tubular cristae and an abundant smooth ER so characteristic of steroid-secreting cells. The fully formed corpus luteum secretes both estrogens and progestins..."
After the first trimester of pregnancy, or if pregnancy does not ensue, the corpus luteum degenerates into a fibrous scar called a corpus albicans.
|
 |
|
< |
|
|
|
|
SIUC / School
of Medicine / Anatomy / David
King
https://histology.siu.edu/erg/ovary.htm
Last updated: 20vMay 2022 / dgk