
Skeletal Tissue (Bone and Cartilage)
INTRODUCTORY COMMENTS
Several varieties of specialized connective tissue together form the skeleton. Appreciating the composition of these tissues is important for understanding the mechanical properties of the skeleton and its susceptibility to various types injury. Furthermore, understanding the processes of normal (and abnormal) skeletal development, of healing after injury, and of degeneration requires some knowledge of the cells which produce these tissues.
Bone and cartilage, like all other connective tissues, consist of cells and extracellular matrix. It is the ground substance of the matrix which is most responsible for the conspicuous differences between bone and cartilage. The ground substance of bone is mineralized, making the bone rigid and strong, but brittle. The ground substance of cartilage is not mineralized but is more like very firm Jello®, making cartilage stiff and incompressible but more flexible and resilient than bone.
Yet, in spite of their solidity, both bone and cartilage are capable of growth. In the case of bone, internal remodelling (essentially, ongoing destruction and renewal) is an active process throughout life.
In both cartilage and bone, the microscopic appearance of ground substance tends to obscure the presence of collagen. However, as in most other connective tissues (other than blood), collagen is a principal fibrous component which confers tensile strength. (The exceptionally flexible cartilage of ears and epiglottis is also heavily invested with elastin.)
CLINICAL NOTE: A balance between osteoclast and osteoblast activity is necessary for a stable calcium level in blood. To this end, osteoclast activity is hormonally regulated, stimulated by parathyroid hormone and inhibited by calcitonin from C-cells of the thyroid.
Clinical relevance of skeletal tissue histology
Broken bones heal by an acceleration of the normally ongoing process of bone remodeling. The break is first patched by formation of new primary (woven) bone and is then gradually reshaped to approximate the bone's original texture and strength.
Initial treatment for broken bones usually involves basic mechanical considerations, stabilizing the structure to prevent further injury. And of course some breaks, such as cervical fracture, immediately raise urgent neurological concerns. But optimizing subsequent repair (which should minimize long-term impairment) requires some considerable understanding of the mechanical properties and healing process of bone and cartilage.
Because we do not have slides of broken and healing bone, the various aspects of bone structure, growth and remodelling must be observed piecemeal in other specimens.
- The basic structure of bone matrix can be seen in the slide labelled "ground bone."
- The cellular elements of bone can be best seen in slides labelled developing cartilage bone and fibrocartilage.
- The process of bone remodelling, similar to process of fracture repair, with osteoblasts laying down new lamellae while nearby osteoclasts are removing previously formed bone, can also be found on slides of developing cartilage bone and fibrocartilage.
Bone which has been remodelled, or which was originally laid down on a neat preexisting bony surface, displays a pattern called lamellar bone. Bone that has been created without such a surface, as in initial repair of a fracture, is less well organized and is described as primary or woven bone.
SLIDE GUIDE: What can be seen, and what should be noticed, on selected slides from the reference set. Each of these slides shows some feature of skeletal tissues. (LUMEN has "Zoomify" slides that are similar to several of these.)
Note: All slides of bone in our slide-set, EXCEPT those labelled "Ground Bone," represent decalcified bone from which the mineral has been removed, while leaving behind cells and organic matrix.]
Ground Bone, x.s.
Gound Bone, l.s.
Decalcified Bone (rabbit femur), x.s. or l.s.
Developing Cartilage Bone
Trachea and lung
Fibrocartilage
Elastic Cartilage
Hyaline Cartilage
Early Dental Cup
Internal Ear
Our "ground bone" slide displays a piece of bone from which all organic material has been removed and the remaining mineral matrix prepared by grinding a small chip of bone into a slab thin enough to transmit light (hence "ground bone," where "ground" is the past-tense of "grind"). The gritty texture which may be observed within most Haversian canals of these specimens is bone dust, an artifact of this preparation by grinding.
The two slides ("x.s." vs. "l.s.") differ only in plane of section -- the cross section ("x.s.") displays concentric lamellae surrounding Haversian canals; the longitudinal section ("l.s.") shows how the lamellae and canals run longitudinally along the long axis of the bone. Both slides display lacunae with interconnecting canaliculi. Since canaliculi are NOT normally visible in decalcified bone, these are our best slides for observing these channels. [Exercise: On a cross section of "ground bone," try to reason out which lamellar systems were formed most recently and which pieces represent the oldest relics of former osteons; see remodelling below.]
The slide labelled "rabbit femur" displays decalcified bone from which the mineral has been removed, leaving behind cells and organic matrix. The section may be either cross section ("x.s.") or longitudinal section ("l.s."). Most features of bone (but not the canaliculi, which are only visible on ground bone) may be found on these slides, which include not only bone but also bone marrow (on the inside) and often periosteum and muscle (on the outside). Active remodelling is apparent on some of these slides (evident by the presence of active osteoblasts and osteoclasts), but the others display quiescent bone. Some of the slides showing active remodelling also display some blue-stained patches within the otherwise-eosinophilic bone; these represent remnants of calcified cartilage which has not yet been replaced by bone.
The slide labelled "developing cartilage bone" displays a longitudinal section through the end of a long bone, at a fairly youthful stage in development when only the shaft (diaphysis) has been mineralized and the end is still cartilagenous. The slide illustrates the process of endochondral ossification (in which cartilage is growing outward toward the end of the bone, while bone tissue is following along behind, invading and replacing the cartilage. In addition to endochondral ossification, you can of course also see hyaline cartilage (the cartilage which faces the articular surface), trabecular bone, cortical bone, bone marrow, muscle, and usually some active osteoblasts and osteoclasts (already remodelling the freshly formed bone). Some of the slides include ends of two bones meeting at a synovial joint, with endochondral ossification occurring in the ends of both long bones.
The slide labelled "fibrocartilage" displays considerably more than fibrocartilage. It appears to be a section through the pubic symphysis, with a more-or-less symmetrical arrangement of bone and muscle on either side of the midline, with the bones bound together by a fibrocartilagenous joint in the medial region of the preparation. Although collagen is present in all cartilage, microscopically visible strands are only apparent in fibrocartilage, where they contribute extra toughness. Active remodelling may be apparent in the bone on this slide (as evidenced by the presence of active osteoblasts and osteoclasts).
Slides of "trachea" and "trachea and esophagus" display the C-shaped rings of cartilage that reinforce the trachea. Cartilage may also be found in bronchial walls in the slide of "lung".
The slides of "hyaline cartilage" and "elastic cartilage" simply display examples of these two tissue types without much context. The source for the hyaline cartilage is not obvious; the elastic cartilage example may be from epiglottis (deducing from the appearance of the associated epithelium). The elastic cartilage slide is stained for elastin (the fine purple strands within the matrix); otherwise it would look just like hyaline cartilage. Hyaline cartilage may be seen on several other slides, notably "developing cartilage bone," "early dental cup," and "trachea."
The slide labelled "early dental cup" displays many features of facial development in a section across the snout of an unidentified animal fetus. If you enjoy microscopy, figuring out this slide should be fun. Symmetry about the midline should help you get oriented. You can then find major anatomical features such as the tongue (with its texture of crisscrossing muscle fibers), the nasal septum (hyaline cartilage), the jaw, and various skull bones. You can then examine the appearance of various bones in various stages of intramembranous or endochondral ossification. You may also find developing muscles, teeth, salivary glands, and skin.
The slide of "internal ear" displays the many elaborate structures of the inner ear, all embedded in bone. This is also the only slide we have in which some of bone has no obvious lamellar pattern, suggesting it represents primary or woven bone that has not yet been replaced by remodelling. If finding and seeing that sounds exciting to you, then enjoy; but you should first become acquainted with the appearance of bone in other specimens (listed above) which are not as complex.
CARTILAGE
The ground substance of cartilage is characterized by a firm solid gel-like matrix (rather like very firm Jello®) reinforced by collagen. Cartilage is relatively stiff and incompressible. However, since cartilage is not mineralized, it is more flexible and resilient than bone.
The characteristic microscopic texture of cartilage, with cells enclosed in individual lacunae (small chambers), reflects the nature of cartilage matrix and its mode of formation. While cartilage is growing, new cartilage matrix is secreted by chondroblasts, which become isolated from one another as the matrix is formed. As more matrix is secreted, these chondroblasts are pushed farther apart. Chondroblasts divide within their lacunae to form "isogenous cell clusters," which then form more matrix and become separated from one another. Once growth ceases, the resting cells are called chondrocytes. Although apparently quiescent, chondrocytes continue to interact at a molecular level with cartilage matrix (e.g., by recycling aggrecan, a major proteoglycan in the matrix.).
Disturbance of matrix maintenance by chondrocytes is implicated osteoarthritis and in Kashin-Beck disease (KBD; see Science 324:1378, 2009).
Since cartilage is avascular, as the cartilage increases in bulk, its cells also grow farther and farther from their source of nutrition. As a direct result, mature cartilage is a relatively inactive tissue with minimal ability to respond to injury. Nevertheless, cartilage remains a living tissue (as does bone): Healthy chondrocytes are necessary to maintain the molecular integrity of the cartilage matrix.
Three forms of cartilage are conventionally recognized:
- Hyaline cartilage, the most common form, is a precursor to developing bone and is also found in synovial joints and as reinforcement for trachea and bronchi.
- Fibrocartilage, reinforced for toughness with collagen fibers, is found in intervertebral discs as well as the pubic symphysis.
- Elastic cartilage, more flexible and springy, is found in ear and epiglottis.
- A fourth type, lipocartilage, has recently been recognized.
Lipocartilage, found in the ears, nose and throat, is based on fat-filled cells called "lipochondrocytes" that allow the tissue to remain soft and springy, "similar to bubbled packaging material." Lipocartilage has only been recently (re)discovered by Raul Ramos et al., after having been first described by Franz von Leydig in 1854. (For a nice summary, see Science 387:136-137, 9 Jan. 2025)
BONE
Perhaps the most surprising attribute of bones, at least for beginners, is that bones are alive. Bones are not just dead, mineralized structural elements. The material of bone is constantly being worked-over (remodelled) through the activities of the cells contained within. And this ongoing activity is an essential part of the body's overall metabolism, maintaining appropriate calcium ion concentrations throughout the body.
The composition and histological organization of bone are well-described in most histology books. (REMINDER: Using this website should not be a substitute for reading a real textbook.) What follows is primarily a guide to observing particular features microscopically.
Most of the significant features of bone can be observed in your reference slides, but not every feature will be apparent on every slide. To properly appreciate each slide, you need to realize whether the bone is still developing (i.e., with sites of endochondral or intramembranous ossification) or mature, and also to determine whether or not active remodelling is visible.
Most mature bones are more-or-less hollow. (Within the hollow is bone marrow.) The outer bone is called cortical or compact bone, while the inner marrow cavity may be criss-crossed by thin strands of bone called trabeculae.

- All mature bone is formed in layers called lamellae.
- Within the lamellae are small spaces, or lacunae, in which osteocytes reside.
- Neighboring lacunae are interconnected by thin channels, or canaliculi.
- In compact bone, the lamellae are organized into sets of concentric rings called osteons or Haversian systems.
- Each osteon includes a central channel, the Haversian canal, which contains a blood vessel.
Historical note: The eponyms "Haversian canal" and "Haversian system" commemorate Clopton Havers, b. 1657.
- The lamellar systems (osteons) of bone are continually being remodelled.
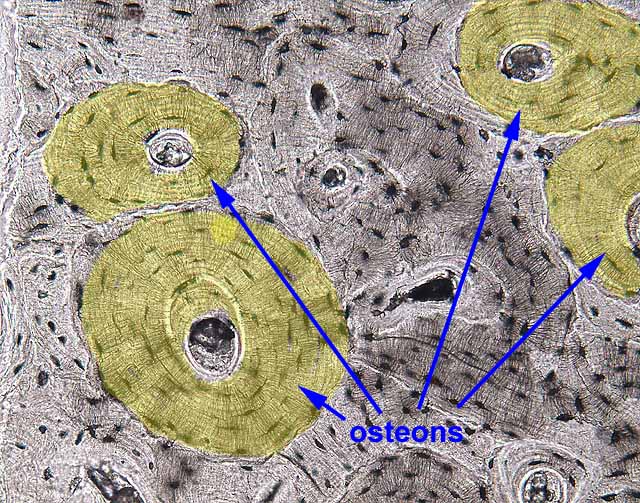
Well-formed Haversian systems are best displayed on our slides of "ground bone." They will not be found in primary, or woven, bone (bone which has not yet undergone remodelling), examples of which may be observed in some of our slides of "internal ear" (in the bony labyrinth around the cochlea).
In any bone with well-developed lamellae, only some of the lamellae will appear as neat concentric Haversian systems (i.e., with a roughly circular outline and a Haversian canal in the center). Other lamellae appear to form only partial osteons, with some portion replaced by the more well-formed lamellae of an adjacent osteon. Still more lamellae appear only as haphazard pieces, with no central canal. This pattern of overlapping osteons and fragmentary lamellar systems provides a visible record of past and recent remodelling.
Exercise: On a cross section of "ground bone," try to reason out which lamellar systems were formed most recently and which pieces represent the oldest relics of former osteons.
Other slides of bone in your reference set ("rabbit femur," "fibrocartilage," "inner ear") represent decalcified tissue, in which details of cells are visible. Collagen in the matrix is left intact by the decalcification process, so the bone matrix appears eosinophilic (i.e., pink in H&E-stained material). Matrix details (especially canaliculi) are less apparent than in ground bone. In mature resting bone, including the slide of "rabbit femur," osteoclasts are rare and osteoblasts are represented only by small, inactive bone-lining cells. Both of these cell types can be readily found in their active states in the slide of "developing cartilage bone," where bone is being actively resorbed and laid down, and also in the slide of "fibrocartilage."
BONE CELLS
- Osteocytes are small, inactive cells, seemingly isolated from one another in individual lacunae. Inconspicuous cell processes extend out through tiny canaliculi and provide gap-junctional contact among neighboring osteocytes as a means of communication and nutrient supply through the otherwise-impermeable bone matrix.
- Osteoblasts (bone-forming cells) are small cuboidal cells, usually found lying adjacent to one another upon lamellae they have just secreted. (A row of osteoblasts can superficially resemble a simple cuboidal epithelium.)
- Osteoblasts lay down new bone lamellae. They are active during bone development and also during bone remodelling.
- Inactive osteoblasts may be called "bone-lining cells."
- Osteoclasts (bone-removing cells) are large cells with multiple nuclei, each one typically sitting alone within a small hollow called a "Howship's lacuna" (the name commemorates John Howship, b. 1781). These lacunae are created by the osteoclasts themselves, as they remove adjacent bone matrix during remodelling.
- Osteoclasts remove preexisting bone. They are active in bone development and also in bone remodelling.
- Osteoclasts are more closely related to macrophages than to osteocytes or osteoblasts. They form a distinct cell population derived from the same precursor cells as macrophages.
These three cell types can be identified not only by their appearance as cells but also by their position in relation to the adjacent bone lamellae:
- Osteocytes are isolated within lacunae, separated from other cells by intervening bone matrix.
- Osteoblasts occur along a surface which parallels the lamellae that they are producing.
- Osteoclasts occur in small hollows (called Howship's lacunae) which they have eroded into the surface. The edges of these hollows typically cut across the underlying lamellae.
CLINICAL NOTE: A balance between osteoclast and osteoblast activity is necessary for a stable calcium level in blood. To this end, osteoclast activity is hormonally regulated, stimulated by parathyroid hormone and inhibited by calcitonin from C-cells of the thyroid.
BONE MARROW
Don't neglect to notice the specialized connective tissues which fill the marrow cavity of many bones, including both hemopoietic (blood-forming) tissue and fat. Bone marrow is not described in this website.
DEVELOPMENT and REMODELLING of Skeletal Tissues
Cartilage formation
Growth of cartilage, from Blue Histology (Copyright Lutz Slomianka 1998-2004)
(The image should be animated, if you watch patiently.)
Growth occurs by two mechanisms.
- Interstitial growth - Chondroblasts within the existing cartilage divide and form small groups of cells, isogenous groups, which produce matrix to become separated from each other by a thin partition of matrix. Interstitial growth occurs mainly in immature cartilage.
- Appositional growth - Mesenchymal cells surrounding the cartilage in the deep part of the perichondrium (or the chondrogenic layer) differentiate into chondroblasts. Appositional growth occurs also in mature cartilage.
In the animation, chondroblast cytoplasm is red, chondroblast nuclei are blue, and cartilage matrix is gray.
Mature cartilage is avascular and relatively permanent. (This is in contrast to mature bone, which is highly vascular and whose extracellular material is continually being remodeled by the cells within, even in adults.) The functional properties of cartilage are derived from relatively stable extracellular materials, not from any ongoing cellular activity. Once cartilage has formed, it is pretty much finished (although it may continue to expand with age and become somewhat less resilient). If adult cartilage is damaged, it may be replaced not by new cartilage but by collagenous scarring. This is especially significant in case of damage to articular cartilages in joints.
Bone develops embryonically in two different ways -- either by intramembranous ossification (directly from mesenchymal condensations) or by endochondral ossification (by transformation of a pre-existing cartilagenous model). Once bones are initially developed, bone continually undergoes a process of remodelling in which old bone is removed and replaced by newly-formed bone.INTRAMEMBRANOUS OSSIFICATION
The simpler mode of bone formation is intramembranous, in which bone forms directly in mesenchyme. This is the pattern for the thin, flat bones of the skull (hence, "intramembranous"). During intramembranous bone formation, ossification occurs in spicules around mesenchymal vessels. Spicules of bone covered by osteoblasts, may be observed in the "dental cup" slide, which shows the snout of a young animal.
ENDOCHONDRAL OSSIFICATION
Most of the embryonic skeleton forms initially as cartilage. During endochondral ossification, osteoblasts and blood vessels must actually invade the cartilage precursor. In long bones (e.g., tibia, humerus), a plate of cartilage remains through adolescence at the junction between epiphysis and diaphysis. This epiphyseal plate is the site of bone elongation, with new cartilage forming on the epiphyseal side of the plate while endochondral ossification extends the diaphysis on the other side of the plate. This arrangement allows the epiphyseal ends of the bone to be solidly calcified (and hence able to bear the brunt of the forces the bone experiences) even while the bone is still growing. Eventually, the ossification front catches up with the chondrogenic front, all of the cartilage is replace by bone, and bone elongation ceases. Timing of this occurrence varies with some predictability from bone to bone and thus contributes to the forensic determination of the age of death from from skeletal remains.
On your reference slide labelled "developing cartilage bone," you can observe endochondral bone formation in the epiphyseal region of a long bone. The entire process, from proliferating chondroblasts, through expanding and hypertrophic cartilage, to mineralization of the cartilage, invasion by blood vessels and osteoblasts,
and remodelling by osteoclasts, can seen sequentially by scanning from the epiphysis toward the diaphysis. A continually advancing growth front has established the appearance of a "layered" structure that is really just a series of points caught at different stages within the same process.
The process of endochondral ossification may also be observed at a synarthrosis in the slide labelled "fibrocartilage."
Bone Remodelling
Living bone is continually being remodelled -- essentially, the matrix is constantly being removed and rebuilt. The process of bone remodelling is extremely active during childhood (maintaining the shape of each bone as the body grows larger) but slows down with advancing age.
Reorganisation and Restoration of Bone, from Blue Histology (Text and GIF copyright Lutz Slomianka 1998-2004)
(The image should be animated, if you watch patiently.)
Changes in the size and shape of bones during the period of growth imply some bone reorganisation. Osteoblasts and osteoclasts constantly deposit and remove bone to adjust its properties to growth-related demands on size and/or changes of tensile and compressive forces.
In the animation, osteoclasts are pink, osteoblasts are blue, and lamellae are gray. Osteocytes (within the lamellae) are not shown.
Although the reorganisation of bone may not result in macroscopically visible changes of bone structure, it continues throughout life to mend damage to bone (e.g. microfractures) and to counteract the wear and tear occurring in bone. Osteoclasts and osteoblasts remain the key players in this process. Osteoclasts "drill" more or less circular tunnels within existing bone matrix. Osteoblasts deposit new lamellae of bone matrix on the walls of these tunnels resulting in the formation of a new Haversian system within the matrix of compact bone. Parts of older Haversian systems, which may remain between the new ones, represent the interstitial lamellae in mature bone. Capillaries and nerves sprout into new Haversian canals.
Restorative activity continues in aged humans (about 2% of the Haversian systems seen in an 84 year old individual contained lamellae that had been formed within 2 weeks prior to death!). However, the Haversian systems tend to be smaller in older individuals and the canals are larger because of slower bone deposition. If these age-related changes in the appearance of the Haversian systems are pronounced they are termed osteopenia or senile osteoporosis. The reduced strength of bone affected by osteoporosis will increase the likelihood of fractures.
The lamellar texture of bone is a direct result of remodeling. Broken bones heal by an acceleration of the normally ongoing process of bone remodeling. The break is first patched by formation of new primary (woven) bone and is then gradually reshaped to approximate the bone's original texture and strength.
CLINICAL NOTE: A balance between osteoclast and osteoblast activity is necessary for a stable calcium level in blood. To this end, osteoclast activity is hormonally regulated, stimulated by parathyroid hormone and inhibited by calcitonin from C-cells of the thyroid.
JOINTS
Joints and associated tissues develop from mesenchyme and may be classified as specialized connective tissue, like other skeletal structures. They are variously composed of fibrocollagenous tissue (principally collagen and fibroblasts) or cartilage. You should appreciate the mechanical differences associated with different types of joint tissue, and be familiar with some specific examples of each type.
Synovial joints have the most specialized structure. The synovial lining is formed from hyaline collagen and synovial cells with no basement membrane. The synovial cavity is thus not lined by a proper epithelium but is more like a highly specialized intercellular space in connective tissue. The synovial fluid is an interstitial fluid, an ultrafiltrate from synovial capillaries, together with highly polymerized hyaluronic acid and glycoprotein secreted by the fibroblast-like synovial cells. This fluid exudes from the synovial membrane and is apparently responsible both for lubricating and nourishing the articular cartilages.
There are normally few white blood cells in the synovial fluid (less than 300 / mm3 vs. about 5000 / mm3 for blood). A significant increase in this number is indicative of joint disease.
The synovium is susceptible to various forms of damage, for example in rheumatoid arthritis.
In our slide sets, some of the slides of developing cartilage bone include the end of an adjacent bone and, between the two, a synovial joint. You can examine various other joints grossly in anatomy lab.
SKELETAL PATHOLOGY
Bone repair (after a break that has been neatly re-set) takes place by a process similar to that of bone remodelling, hopefully resulting freshly-formed bone that eventually is at least as strong as the original. Details of skeletal pathology should be available in any pathology text (e.g., Robbin's Pathological Basis of Disease).
Comments and questions: dgking@siu.edu
SIUC / School
of Medicine / Anatomy / David
King
https://histology.siu.edu/ssb/skeleton.htm
Last updated: 9 February 2025 / dgk