|
Nervous Tissue
|
Neurons and Support Cells
Please note that this guide is intended to complement, NOT to replace, textbook readings.
- Text Reference: Kandel's Principles of Neural Science.
- Introductory comments / How to read this page.
- NERVE CELLS
- SUPPORT TISSUE
- SOME EXAMPLES of nervous tissue
- PERIPHERAL NERVES
- Composition and appearance of peripheral nerves
- Sensory and motor nerve endings
- For organs of special sense, see:
- CENTRAL NERVOUS SYSTEM
- Nervous tissue pathology
- SAQ Self Assessment practice Questions
- Link to virtual slides at LUMEN (Loyola University Medical Education Network), click on "Part 6: Neural Tissue."
- Cells-R-Us (illustrated text of a public lecture on the brain, by the author of this website)
- Image index
Histology textbooks are NOT recommended for the study of nervous tissue. Rather than emphasizing features important for understanding nervous tissue function, most histology textbooks begin with relatively insignificant, and often misleading, details. (For example, many histology texts classify dorsal root ganglion neurons as "pseudo-unipolar," which is accurate but useless, since that that category is never mentioned again. More significantly, the distal branch of such a cell's axon is not infrequently referred to as a "dendrite," simply because it conducts toward the cell body. But by all generally recognized criteria, this process is a sensory axon.)
RECOMMENDED are selected chapters in Kandel's Principles of Neural Science.
Kandel's classic text is remarkable. The extended table of contents can be read, just as if it were a "capsule" textbook. In about two dozen pages following immediately after the chapter listing, all of the subheadings from every chapter are presented, each as a complete sentence. This extended table of contents offers a concise summary of major ideas. Your study through this entire unit can be usefully guided by this summary. You should, at a minimum, be fluent in the vocabulary of this summary so that every sentence here is meaningful to you.
Senior author Eric Kandel shared the 2000 Nobel Prize for Physiology and Medicine for work on "signal transduction in the nervous system" (Kandel's Nobel Prize lecture; Kandel's Nobel banquet speech; Kandel extended biography).
INTRODUCTORY COMMENTS
Please note that this guide is intended to complement, NOT to replace, textbook readings (i.e., Kandel et al.).
If you feel intimidated by heavy textbooks, you might appreciate a collection of 2-minute mini-lectures at NEUROSCIENTIFICALLYCHALLENGED.com.The study of nervous tissue presents extraordinary challenges. Historically, other tissues were fairly well understood decades before science acquired a basic appreciation for the cellular composition of nervous tissue. One reason is that routine histological preparations do not enable proper visualization of any nerve cell type. Significance for study: Do not expect microscopic examination of nervous system specimens to yield satisfying observations, at least not without an exceptional effort to understand why things look the way they do.
☞ An excellent New York Times article, with video, describes in detail how brain tissue is processed for histology in a modern pathology laboratory. (This article [Sept. 9, 2025] was written in the context of diagnosing chronic traumatic encephalopathy, C.T.E.The Neuron Doctrine
Modern understanding of nervous tissue (i.e., since the early 20th century) is summarized by the four principles of The Neuron Doctrine (quotations below are from Eric Kandel's 2006 autobiography In Search of Memory, pp. 65-66):
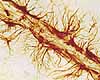
Neuron preparation by Ramón y Cajal Historical note: These four principles were introduced by Santiago Ramón y Cajal, the most famous pioneer in the descriptive anatomy of nerve cells. Cajal's 1906 Nobel Prize lecture includes some elegant images of nerve cells in spinal cord and in cerebellar and cerebral cortex. (For curiosity, see "Milestones in Neuroscience Research," a lengthy list covering several thousand years.)
- Cellularity: "The nerve cell is the fundamental structural and functional element of the brain."
- Synaptic communication: "The terminals of one neuron's axon communicate with the dendrites of another neuron only at specialized sites, later named 'synapses' by Sherrington."
- Connection specificity: "Neurons do not form connections indiscriminately. Rather each nerve cell forms synapses and communicates with certain nerve cells and not with others."
- Dynamic polarization: "Signals in a neural circuit travel in only one direction... Information flows, from the dendrites of a given nerve cell to the cell body [then] along the axon to the presynaptic terminals and then across the synaptic cleft to the dendrites of the next cell, and so on."
Axons, dendrites and synapses -- the most significant features of nerve cells -- cannot be readily seen without specialized techniques such as those used by Cajal. (The spaces between nerve cell bodies are filled with a feltwork of these axonal and dendritic processes, called neuropil, which also includes glial cell processes.)
No other tissue in the body is characterized by cells whose cytoplasmic processes reach out for vast distances away from the cells' nuclei. This reality puts a special burden on the student as well as the researcher. You cannot simply look at a slide or micrograph, not even an electron micrograph, and truly "see" the most interesting features of the nerve cell.
Nerve cell processes are quite thin, often less than a micron (1µm) in diameter. However, the length of axons and dendrites is wondrously great, far greater than ordinary cellular dimensions. Dendrites may extend several millimeters away from the cell body, into a volume the size of a pea. Axon length commonly extends for several centimeters and may exceed a meter for many sensory and motor axons.
As a simple consequence of this cellular geometry, the cell body of a neuron may comprise less than one percent of the cell's total volume. From this, you may correctly infer that the bulk of nervous tissue consists of nerve cell axons and dendrites rather than nerve cell bodies.
The study of neuroanatomy consists largely of understanding the routes travelled by nerve cell axons.
Unfortunately, the organization of neural processes, most particularly the full length of axons and dendrites and the synaptic interactions between them, can seldom be visualized directly.
In most other tissues of the body, what you can see in the microscope is directly informative. Consider skin, where a routine section of epidermis reveals almost everything interesting about the size, shape and growth sequence of epidermal cells. Electron microscopy of similar specimens simply adds more finely resolved detail. But making any sense at all of nervous tissue requires that you "see" with concepts acquired over decades of research using many special techniques.
Historical note: One of the first challenges for early-modern neuroanatomy consisted of mapping long-distance connections between regions. Since it was (and remains) utterly impossible to trace individual axons visually over their entire length, other techniques of "experimental neurology" were needed.
One of the principal methods for nervous system mapping depended on cellular responses to injury following lesion of a region in the nervous system of an experimental animal.
One of these responses is "retrograde degeneration," also known as the "axon reaction" (described in some detail by Franz Nissl in 1894), whereby damage to an axon leads (after a suitable time interval) to alterations in the associated cell body. By painstakingly searching throughout the brain for cell bodies displaying the axon reaction following such a lesion, researchers could discover (at cost to many experimental animals) the source(s) for axons passing into or through the lesioned area.
Similarly, signs of "anterograde degeneration" or "Wallerian degeneration" (named after Augustus Waller, b. 1816), whereby distal portions of a damaged nerve fiber degenerate (again after a suitable time interval) could be used to find the destination(s) of axons originating in or passing through the lesioned area (again, at cost to many experimental animals).
When you examine microscope slides or micrographs of nervous tissue, patterns of functional connection cannot usually be seen. Nevertheless, what you can observe should be interpreted in terms of neuronal functions and connectivity, including unseen axons, dendrites and synapses as well as associated supporting cells.
Thus your job for comprehending nervous tissue is not just to look-and-learn, but to think rather deeply, to fit many different views and facts together. Most of the listed vocabulary terms for neuronal and glial structures are well defined in standard textbooks. You just have to make sense of it all.
How to read this page. This page is much longer than most other pages at this histology website, presumably because its author did his doctoral dissertation on nerve cells. You might read this page straight down, from top to bottom. But it is written with hyperlinks to facilitate browsing. You might more profitably check out each link, at least if it suggests a question in your mind, and use your browser's "back" arrow to return. And return repeatedly to the outline at the top of this page to choose the topic that most closely engages your current curiosity.
BASIC STRUCTURE OF NERVE CELLS ("NEURONS")
"Swiftly the brain becomes an enchanted loom, where millions of flashing shuttles weave a dissolving pattern -- always a meaningful pattern -- though never an abiding one" (Sherrington, 1940).
Nerve cells comprise the "enchanted loom" that is our brain.
Here are three absolutely wonderful facts about nerve cells.
- The first and most wonderful fact is that, working together, nerve cells can perceive and think and dream. They are us. This is magic of the highest sort. (See my essay Cells-R-Us for informal discussion.)
- The second wonderful fact is that nerve cells are much like other cells. Each is essentially a bag of water, surrounded by a fatty membrane and containing an assortment of molecules. There seems to be nothing about individual nerve cells that cannot be explained, at least in principle, by basic chemistry and biology.
- The third wonderful fact is that each nerve cell has a truly magnificent shape. Somehow, this third fact bridges the gap between the mystery that is our mind and the chemistry that is our cells. Somewhere in the shape of nerve cells, in the complexity of connections among billions of such cells, and in the intricate pattern of activity that plays upon those cells, our "self" emerges.
Nerve cells come in extreme variety. In every region of the brain are several different nerve cell types, each distinguished by its own characteristic soma size, dendritic shape, source of synaptic input, destination of axonal output, and chemistry (more below). Occasional nerve cell types may have characters which depart from the the typical description presented below.
Because of this immense variety of nerve cell types, there is no "one-size-fits-all" description. So textbook descriptions of nerve cells tend to present overwhelmingly abundant detail. Although many details of nerve cell shape and connectivity are usually insignificant for clinical practice, they can be quite beautiful. (Ramón y Cajal famously referred to nerve cells as "the mysterious butterflies of the soul.") And some details are essential for understanding research on brain function. It is also often necessary to learn some "irrelevant" detail in order to understand the particular examples used to demonstrate basic functional principles.
*** Most of the following generalities have exceptions. ***
Every nerve cell has three distinctive portions: a cell body, one axon, and several dendrites.
- The cell body of a nerve cell (also called the perikaryon or soma [plural somata]) is basically a cell nucleus surrounded by cytoplasm. (Perikaryon means "around the nucleus.") Nerve cell bodies look more or less like other body cells, although they do have certain distinguishing features:
Extending out from each nerve cell body are long cytoplasmic processes: one axon and several dendrites. (These processes usually cannot be distinguished in routine histological preparation.)
- The nucleus of a nerve cell is typically large, round and euchromatic with a single prominent nucleolus (more below).
- Cytoplasm of a typical nerve cell body is abundantly supplied with masses of rough endoplasmic reticulum (traditionally called Nissl bodies, named for Franz Nissl, b. 1860), numerous Golgi bodies (named for Camillo Golgi, b. 1843), lots of smooth endoplasmic reticulum, and many mitochondria. Extensive cytoskeletal elements (microtubules and various filaments) extend from cell bodies into neural processes, where they provide the essential framework for ongoing maintenance of axons and dendrites.
Historical note: One of the pioneers in the study of the neuronal cytoskeleton was Sigmund Freud, during his student research into comparative neuroanatomy of crayfish. For more on cytoskeletal research, see The Cytoskeleton of Nerve Cells in Historical Perspective, by E. Frixione, IBRO History of Neuroscience, 2006. This essay also describes the role played by the neuronal cytoskeleton in development of the neuron doctrine, including significant contributions by Ramón y Cajal- A typical nerve cell body contains only a tiny fraction of the total cell volume; the rest is contained in the axon and dendrites.
- The spaces between nerve cell bodies is filled with a feltwork of axonal and dendritic processes, called neuropil, as well as glial cells and their processes.
- The axon is a process which is specialized for conducting signals over long distances from one nerve cell to another.
Each nerve cell has one and only one axon, although that axon may branch extensively.
- Typical axons have relatively few branches, except near their terminal ends.
- The terminal branches of an axon make synaptic contacts onto other nerve cells (or with peripheral effectors, i.e., muscles and glands).
- The diameter of an axon is uniform along most of its entire length.
[Recent research (Science, 05 Dec 2024, 386:1084-1085) has suggested that unmyelinated axons might, under some conditions, assume a "string of pearls" shape; the evidence has met with skepticism as possibly representing a preparation artefact.]- Nerve signals travel along axons away from the cell body and toward synapses at the axonal terminal.
- Axonal nerve signals, called action potentials, are initiated at the axon hillock, the site where an axon arises from the cell body.
- Action potentials are active, all-or-nothing signals which do not decline in strength as they travel along the axon.
- Axons may or may not be myelinated. Myelination increases the speed of signal propagation by enabling saltatory conduction.
- On a cellular scale, the dimensions of an axon may be enormous: axonal length may exceed ordinary cellular dimensions by several orders of magnitude .
- Many axons reach a length of several centimeters. Axons in peripheral nerves can be over a meter long. (This is in contrast to dimensions on the order of 5 to 50 micrometers for most non-neural cells.)
- The volume of an axon may be hundreds of times greater than the volume of an ordinary cell.
- The membrane surface area of an axon may be hundreds of times greater than that of an ordinary cell.
- Maintaining such vast material requires impressive metabolic machinery in the cell body as well as efficient mechanisms for axoplasmic transport up and down the axon.
Clinical note: Certain viruses (such as the rabies virus or Herpes zoster) coopt axonal transport mechanisms to migrate from the periphery into neuron cell bodies. Shingles results when herpes is transported from its "hiding place" in dorsal root ganglion cells back out into the periphery.- Damaged axons undergo "Wallerian degeneration" (named after Augustus Waller, b. 1916), eventually dying and disappearing distal to the site of injury.
Clinical notes: If a peripheral nerve is severed or crushed, the proximal portion of each axon, which is still connected to its cell body, can regrow distally from the site of injury. The "growth cone," leading this outgrowth, follows rows of Schwann cells -- provided that the damaged parts of the nerve remain well aligned.Following traumatic amputation, surgical repair calls for reattachment of the cut or torn ends of the epineurium to help assure alignment for axon regrowth. Restoration of function requires axon regrowth -- a slow process of about an inch per month -- which depends on the rate of axoplasmic transport of material from the cell body. (See, e.g., A. Höke, 2011, J. Clin. Invest. 121(11): 4231-4234.)
Dendrites are processes which are specialized for receiving and integrating signals from other nerve cells. ("Integrating" is the jargon term for what dendrites do when they combine synaptic input from several sources.)
Neuron preparation by Ramón y Cajal
A nerve cell typically has several dendrites, each with numerous branches. (The word "dendrite" derives from a word meaning "tree.") The diameter of dendrites typically decreases away from the cell body, so that dendrites taper gradually to fine twigs.
- Dendrites typically receive synaptic contacts from axons of many other nerve cells. Synapses often occur on tiny dendritic spines.
- Synapses onto a single nerve cell's dendritic branches may number in the thousands, or even hundreds of thousands.
- Dendritic nerve signals, called synaptic potentials, arise at synapses.
- Synaptic potentials are conducted passively along dendrites toward the cell body.
- Synaptic potentials fade (i.e., decrease in amplitude) with distance from their originating synapse.
- Dendrites are not myelinated, although some large dendrites may support action potentials.
- Size and shape of dendritic spines influence synaptic strength. Plasticity of dendritic spine morphology is implicated in memory.
- Synapses are points of contact between nerve cells (usually between axon terminals and dendrites), where signals are transmitted from one cell to another. (Synapses can seldom be resolved without electron microscopy.)
Historical note: "Giant synapses," such as the "calyces of Held" in the brainstem auditory pathway, are an exception. These were first reported by Hans Held in 1893 and subsequently described in detail by Ramón y Cajal using light microscopy. The "calyx of Held" has since become a model system for studying synaptic function in mammalian nervous systems.
- Neurotransmission is usually chemical, based on small molecules called neurotransmitters, secreted by one cell and binding to another.
- Neurotransmitter binding typically causes changes in membrane ion conductance, thereby locally altering dendritic membrane potential.
- Such local synaptic potentials spread passively, to be integrated with other synaptic potentials, typically at the axon hillock.
- (Neurotransmission can also be electrical, with ions passing directly from one cell into another via gap junctions.)
- Each synapse has a presynaptic side: an axon terminal, from which a neurotransmitter is released.
- Neurotransmitter is usually stored in synaptic vesicles within the presynaptic terminal.
- Neurotransmitter is released in response to changes in membrane potential associated with arrival of action potentials.
- Each synapse has a postsynaptic side: a dendrite or a nerve cell body, where the membrane is specialized to respond to the binding of neurotransmitter molecules, e.g. by altering membrane ion conductance.
- Size and shape of dendritic spines influence synaptic strength. Plasticity of dendritic spine morphology is implicated in memory.
- Neuromodulation involves transmitters with more diffuse function (i.e., less localized to specific synapses, less-clearly associated with discrete histological features). [More at Wikipedia.]
*** Most of the preceding generalities have exceptions. ***
BASIC ELECTRICAL FUNCTION of nerve cell membranes
Note: Most of this entire histology website focuses on cell and tissue structure (i.e., features which can be observed with a microscope). My presumption in creating this website has been that most details of cell function will be covered by courses in physiology and/or cell biology. However, because details of nerve cell shape are so closely associated with the electrical behavior of nerve cell membranes, this section offers an outline sketch (without illustrations) of how nerve cells manipulate localized changes in cell membrane potentials in order to carry out their functions of processing and transmitting information. (Readers not yet familiar with membrane electrochemistry are encouraged to read this section from top to bottom at least twice, to see how the various ideas fit together.)Skip this section (next section is myelin).
Caveat: This section is extremely elementary and highly simplified. For an alternative (and illustrated) presentation, see links immediately below to Chapters 6 and 7 in Principles of Neural Science, 5th ed., by Kandel et al., 2013:
- Membrane Potential and the Passive Electrical Properties of the Neuron (Chapter 6, Kandel et al., 5th ed.).
- Propagated Signaling: The Action Potential (Chapter 7, Kandel et al., 5th ed.).
Core ideas:
- Nerve cells integrate and transmit information by manipulating local changes in membrane potential.
- Local changes in membrane potential are regulated by ion channels in nerve cell membranes.
- The conductance of specific ion channels may change in response to chemical and electrical signals.
- Synaptic potentials result from neurotransmitters acting on specific ion channels in postsynaptic membranes.
- Action potentials result from stimulation of voltage-dependent sodium ion (Na+) channels in axonal membranes.
- Voltage-dependent Na+ channels characterize axons (not most dendrites), generally beginning with the axon hillock.
- Along myelinated axons, voltage-dependent Na+ channels are restricted to nodes of Ranvier.
- Voltage-dependent Na+ channels also characterize muscle fibers.
- Clinical relevance: Many drugs and toxins can modify the behavior of ion channels.
Membrane Potentials:
- equilibrium potentials and the Nernst equation;
- resting potential and the Goldman equation;
- synaptic potentials;
- action potentials.
(Note that there exist abundant internet resources to facilitate more-detailed understanding neuronal membrane electrochemistry, e.g., Michigan State University, Khan Academy.)
Ion-specific EQUILIBRIUM POTENTIALS and the NERNST EQUATION
- The concentrations of ions such as Na+ and K+ inside every cell differ from those outside.
- An ion's equilibrium potential (also called the Nernst potential or the "reversal potential") is the voltage across a membrane which would balance the tendency of that ion to diffuse across the membrane, following its concentration gradient.
- For any ion whose concentration differs across a membrane, that ion's equilibrium potential is given by the Nernst equation.
- Variables in the Nernst equation are the ion's concentrations on each side of the membrane.
- For several ions whose concentrations differ across a membrane, each ion will have its own specific equilibrium potential.
- (The Nernst equation itself is based on fundamental thermodynamic principles, but the equilibrium potential for an ion can also be determined experimentally.)
RESTING MEMBRANE POTENTIAL and the GOLDMAN EQUATION
- Every cell has an electrical potential difference (i.e., voltage) across its membrane, called the cell's resting potential.
- Resting potentials originate from differing ion concentrations between the inside and the outside of a cell, together with variation in specific ion conductances across the membrane. (Conductance quantifies how freely an ion can pass across the membrane.)
- At rest, the following conditions usually apply:
- Sodium ion (Na+) concentration is higher outside the cell.
- Na+ conductance is relatively low.
- Potassium ion (K+) concentration is higher inside the cell.
- K+ conductance is relatively high.
- (Calcium ions [Ca++], chloride ions [Cl-], and organic anions also contribute to membrane potentials.)
- Local changes in ion conductance can cause localized departures from the resting potential.
- For any given ion, the membrane's resting potential will generally differ from that ion's equilibrium potential.
- Therefore the resting potential is a dynamic equilibrium for multiple ions, each of which has its own particular equilibrium potential.
- Under such conditions, the resting potential entails continual movement of ions across the membrane.
- To compensate for this ongoing ion diffusion, appropriate ion concentrations inside a cell are maintained by the action of sodium-potassium pumps, with ATP providing energy to move ions against their concentration gradients.
- The resting potential for any cell membrane is given by the Goldman equation.
- Variables in the Goldman equation include not only each ion's concentration on each side of the membrane but also each ion's conductance across the membrane (i.e., how readily the ion can pass through the membrane via ion channels).
- (Like the Nernst equation, the Goldman equation is based on fundamental thermodynamic principles.)
- A normal cellular resting potential is dominated by K+ conductance, which at rest is substantially greater than Na+ conductance.
- Therefore, a cell's resting potential generally lies close to the K+ equilibrium potential.
- Any ion not at equilibrium at the resting potential will diffuse down its electro-chemical gradient.
- Therefore, the resting potential would decay to a stable state (where all ions have the same equilibrium potential) unless ion concentrations were maintained by sodium-potassium pumps which counteract diffusion while consuming energy.
Local changes in ion conductance can cause localized departures from the resting potential. ION CHANNELS and ion conductance
- Ion movement across a membrane occurs via dedicated, ion-specific ion channels.
- Ion channels consist of trans-membrane protein complexes.
- Ion movement through ion channels follows that ion's electro-chemical gradient.
- The conductance for an ion is determined both by how freely the ion can pass through an individual channel and by the number of channels.
- The conductance at particular membrane locations may vary in response to chemical or electrical stimuli.
- Various ion channels in postsynaptic membranes respond to neurotransmitters by altering ion conductance to produce excitatory or inhibitory synaptic potentials.
- Voltage-dependent sodium channels in axonal membranes respond to depolarizing membrane potential change by initiating action potentials.
- When "open," an ion channel allows relatively free passage.
- When "closed," an ion channel may still be "leaky," (i.e., always allowing some movement).
- Na+ and K+ each have their own dedicated channels.
- Internal Na+ and K+ concentrations are maintained by active (energy-requiring) sodium-potassium pumps.
- Clinical relevance: Ion channels are targets not only for neurotransmitters but also for many drugs and toxins, which can alter their properties.
SYNAPTIC POTENTIALS
- Synaptic potentials are local departures from resting potential, initiated at sites of synaptic contact between nerve cells.
- Arrival of an action potential at an axon terminal triggers release of neurotransmitter.
- Neurotransmitter diffuses across the synaptic cleft and binds to transmitter receptors associated with specific ion channels in the postsynaptic membrane.
- Transmitter binding initiates processes which alter ion conductance through the associated ion channels.
- Altered ion conductance at a postsynaptic membrane causes a localized change in membrane potential at that site.
- Local postsynaptic membrane shifts may be "excitatory" (depolarizing the membrane away from its resting state) or "inhibitory" (resisting depolarization).
- Localized postsynaptic membrane potential changes cause currents which flow away from the postsynaptic site, with the result that the postsynaptic potential spreads passively from the postsynaptic site, decreasing in amplitude over distance.
- Synaptic potentials are variable in magnitude, depending on variation in any of the factors which produce them.
- Spreading synaptic potentials from multiple synaptic sites interact with one another.
- At any given site on a dendrite or cell body, interacting synaptic potentials shift the membrane potential away from its resting state.
- If interacting synaptic potentials exceed a threshhold at an "action potential initiation site" (a site which contains voltage-dependent sodium channels, generally located at or near an axon hillock), an action potential will be initiated.
ACTION POTENTIALS and the Hodgkin-Huxley equations.
- Action potentials are brief, actively-propagating, all-or-nothing waves of depolarizing membrane potential. Action potentials travel without decrement away from the site of their initiation.
- The ability to sustain action potentials is characteristic of axons and striated muscle fibers.
- The measured changes during an action potential are:
- A sudden and substantial increase in Na+ conductance, such that Na+ conductance becomes greater than K+ conductance.
- A consequent rapid depolarizing shift in membrane potential, toward the Na+ equilibrium potential.
- A subsequent increase in K+ conductance, such that K+ is again greater than Na+.
- A consequent rapid repolarization of the membrane, as the membrane potential again approaches the K+ equilibrium potential.
- A restoration of normal, resting conductances for Na+ and K+, and hence of the normal resting potential.
- Thus an action potential consists of a rapid local depolarization of the axon membrane, followed almost immediately by a rapid repolarization, and further followed by a brief "refractory period" during which that particular site will not generate another action potential.
- The molecules which respond initially are called "voltage-dependent sodium channels," or "voltage-gated sodium channels."
- An action potential is triggered by a depolarizing change in membrane potential which exceeds a threshhold for activating voltage-dependent sodium channels.
- Normally, action potentials are initiated at the most proximal site which contains voltage-dependent sodium channels, usually near an axon's origin at the cell body, a site which is sometimes visibly differentiated as an "axon hillock."
- At such a site, the initiating potential change is generally the summation of many depolarizing synaptic potentials.
- Once an action potential is initiated, the spread of depolarizing potential away from the initiation site triggers a similar response from adjacent voltage-dependent sodium channels.
- In that way, an action potential actively propagates itself away from its initiation site.
- The recovery time to restore the resting potential at a site that has just experienced an action potential prevents the action potential from propagating "backwards" in the direction from which it had originated.
- Along myelinated axons, voltage-dependent Na+ channels are restricted to nodes of Ranvier.
- The presence of myelin increases the resistance and decreases capacitance of the axonal membrane, thereby allowing action potentials to "jump" from one node of Ranvier to the next. This phenomenon, known as saltatory conduction, greatly increases the speed of action potential propagation.
- (The changes in ion conductance, and hence in membrane potential, during an action potential are described quantitatively by the Hodgkin-Huxley equations; these equations comprise an empirical model, based on experimental measurements taken during action potentials.)
Reiterated caveat: The information in the section above is extremely elementary and highly simplified. For an alternative, illustrated presentation, see links below to Chapters 6 and 7 in Principles of Neural Science, 5th ed., by Kandel et al., 2013:
- Membrane Potential and the Passive Electrical Properties of the Neuron (Chapter 6, Kandel et al., 5th ed.).
- Propagated Signaling: The Action Potential (Chapter 7, Kandel et al., 5th ed.).
MYELIN

Myelin is a fatty covering which envelops many axons.
Myelin enables saltatory conduction, permitting action potentials to be propagated at a much greater velocity than is possible along unmyelinated axons.
- Axons with myelin are called myelinated axons.
- Most myelinated axons are fairly large, ranging from 1µm up to 10µm in diameter (not counting the myelin). To put this size in perspective, the diameter of a large axon may be greater than that of a capillary.
- Axons without myelin are called, logically enough, unmyelinated axons.
- Unmyelinated axons are usually quite small, less than 1µm in diameter.
Myelin is formed by support cells (Schwann cells in the peripheral nerve system, oligodendroglia in the CNS) wrapping around the axons. Myelin is not part of, nor produced by, the nerve cell whose axon it envelops.
In peripheral nerves, myelin consists of Schwann cell membrane wrapped around and around an axon. Most of the Schwann cell cytoplasm lies alongside the myelin wrapping. (See oligodendroglia for myelination of CNS axons.)
To visualize myelin formation:
- Imagine that a Schwann cell is a pillow with the pillowcase representing Schwann cell membrane and the pillow's stuffing representing Schwann cell nucleus and cytoplasm.
- Next imagine a broomstick (representing the axon) lying across one end of the pillow.
- Now roll the broomstick up in the pillow, wrapping the pillowcase tightly around and around the broomstick while squeezing the pillow's stuffing into one end.
- The tight wrappings of pillowcase now represent the myelin, while the remaining pillow with stuffing represents the Schwann cell body with nucleus and cytoplasm.
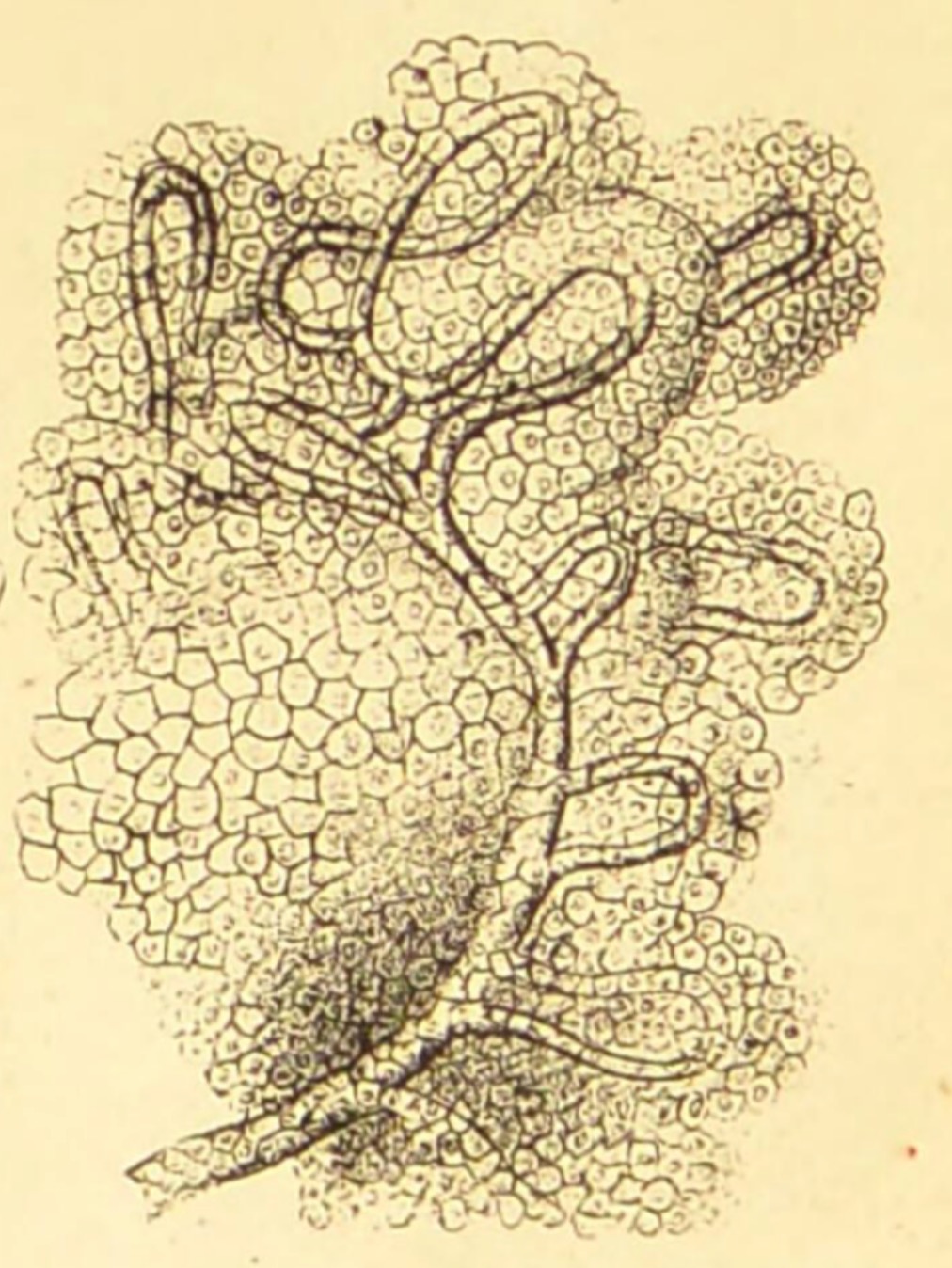
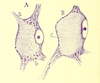
Myelination of a peripheral axon
Animation from Blue Histology, copyright Lutz Slomianka 1998-2004(The image should be animated, if you watch patiently.)
A Schwann cell is illustrated with brown cytoplasm.
The blue oval is the Schwann cell's nucleus.
Observe that as the growing Schwann cell spirals inward around the axon, it wraps its membrane into layers of myelin.
The myelin of one Schwann cell wraps about one to two millimeters along an axon. To myelinate the entire length of the axon, many of these Schwann cell wrappings line up end-to-end along the axon.
The points between segments of myelin are called nodes of Ranvier (named after Louis Ranvier, b. 1835). The stretch of axon between nodes is called an internode.
DETAIL: Cytoplasm around a Schwann cell's nucleus is connected -- by narrow channels which spiral inward beside the nodes of Ranvier at either end of an internode -- to the cytoplasm alongside the axon. Similar channels within the internode are called Schmidt-Lanterman clefts (commemorating H.D. Schmidt, b. 1823, and A.J. Lanterman, b. 1845).NOTE:Many details of myelin cannot be well-appreciated by light microscopy. For electron micrographs of myelin in peripheral nerves, see the online Electron Microscopic Atlas of cells, tissues, and organs (the text is in German, but most figure labels can be deciphered fairly easily).
SALTATORY CONDUCTION

The spacing of nodes of Ranvier is critical for propagation of action potentials. Along myelinated axons, action potentials are regenerated only at the nodes. Myelin provides insulation -- and, more importantly, decreased capacitance -- so that the ionic currents at one node can flow efficiently (and quickly) to the next node. This is called saltatory conduction (saltation = jump). In contrast, action potentials propagating along unmyelinated axons are regenerated at each point along the way, a much slower process. [A hydrodynamic metaphor for saltatory conduction may offer a somewhat intuitive explanation.]
Clinical note, multiple sclerosis: Because myelinated axons have voltage-dependent sodium channels only at nodes of Ranvier, even localized sites of demyelination, such as those which occur in multiple sclerosis, can effectively prevent the propagation of action potentials across the lesion.
Clinical note, local anesthesia: As long as myelin is intact, local currents generated by an action potential at one node of Ranvier are generally sufficient to depolarize axonal membrane two or three nodes away. Local anesthesics, which block action potentials but do not prevent current flow, must therefore be distributed across several nodes (several millimeters) in order to produce effective anesthesia.
RECOGNIZING NERVE CELLS in histological preparations.
Although axons reach into all parts of the body, the vast majority of nerve cell bodies occur in the central nervous system (brain and spinal cord), in those regions described as gray matter. Relatively few nerve cell bodies occur peripherally, in the ganglia (small clusters of nerve cells) of sympathetic and parasympathetic nervous systems.
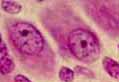
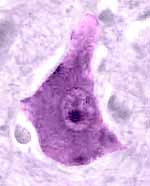
Wherever they occur, most nerve cell bodies have a distinctive appearance.
- Nuclei of nerve cells are large, round and euchromatic with a single prominent nucleolus. Because of this distinctive nuclear appearance, neurons are sometimes described as having "owl-eye" nuclei or "fried-egg" nuclei.
- Cytoplasm of all but the smallest nerve cell bodies is substantial and conspicuously basophilic, containing characteristic basophilic masses of rough endoplasmic reticulum that are traditionally called Nissl bodies (commemorating Franz Nissl, b. 1860).
These features of nerve cell bodies are related to the heavy metabolic demands imposed by maintaining extensive cytoplasmic processes (i.e., axons and dendrites). They are exaggerated (i.e., bigger nuclei, more cytoplasm) in those nerve cells which have the longest, largest diameter axons.
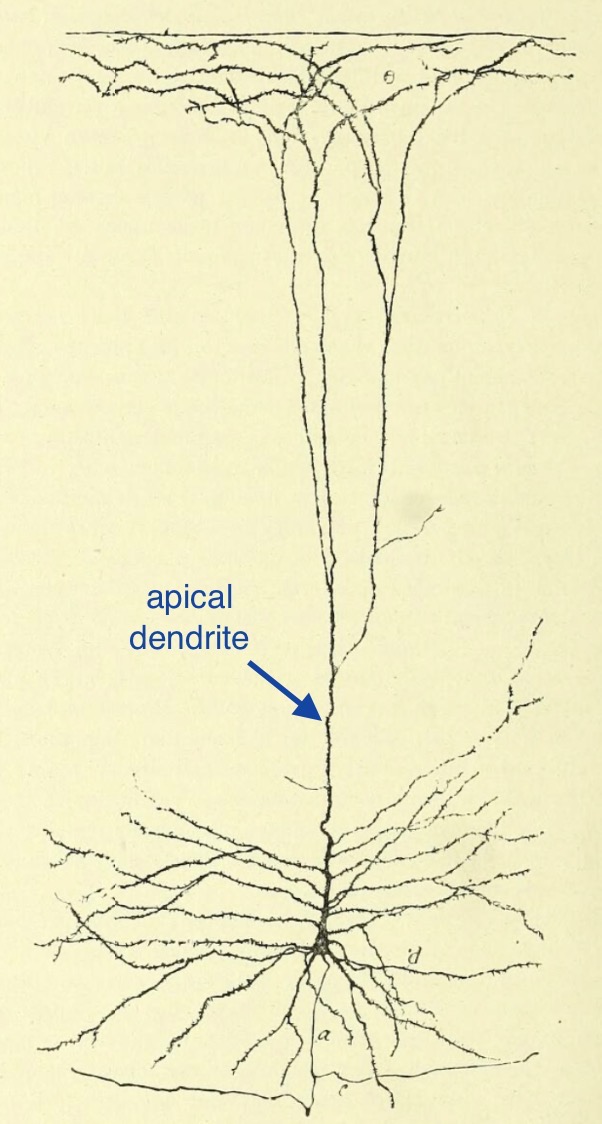
Nerve cells with the most extremely long, large diameter axons -- such as pyramidal cells of motor cortex and motor neurons of spinal cord -- are often illustrated as "typical" neurons simply because they are big and hence especially easy to visualize. Cerebellar Purkinje cells comprise another "popular" type of nerve cell, also large but with a huge dendritic tree rather than an especially long axon.

Neuron preparation by Ramón y Cajal Special stains, like the silver-based Golgi stain, can reveal entire neurons or glial cells (at least as much as fits within the thickness of a single section) by impregnating them with opaque silver salts. But this technique yields elegant results only by suppressing any staining of most neighboring cells, so neurons appear in splendid isolation when their essence is one of complex interaction. Similarly, electron microscopy can display elegant synapses, but the narrow view offers few clues about the cells to which the pre- and post-synaptic profiles belong.
Historical note: The Golgi stain was discovered by Camillo Golgi and was famously exploited by Santiago Ramón y Cajal to develop the Neuron Doctrine.
Sections of central nervous tissue routinely show neuron cell bodies surrounded by a finely-textured fibrous material often called neuropil (which should not be confused with connective tissue). This feltwork consists of axons and dendrites (and glial processes), with all the comings and goings that these processes entail. Individual axons and dendrites can be distinguished only in fortuitous sections, and then only for a short length. The so-called "molecular" layers of cerebral and cerebellar cortex consist of neuropil containing relatively few cell bodies (most of the cell bodies lie in deeper layers).
Note that a common artefact, resulting from tissue shrinkage, is for a clear "halo" to appear around cell bodies and blood vessels. Although the presence of such halos can be misleading (there is no such space in intact, living nervous tissue), this consistent artefact serves to highlight or emphasize the locations for these structures.
SUPPORT CELLS of nervous tissue.
Schwann cells are support cells in peripheral nerves (named after Theodor Schwann, b. 1810).
- Schwann cells form the myelin around myelinated peripheral axons.
- Schwann cells also envelop unmyelinated axons, but without the close, dense membrane wrapping which characterizes myelin.
Clinical note: When a peripheral nerve is crushed or severed, the distal portion of each axon undergoes Wallerian degeneration. However, Schwann cells distal to the injury may remain intact (although their myelin does degenerate); the linear arrangement of Schwann cells can then serve to guide axon regrowth during recovery.Many of the small, heterochromatic nuclei that can be seen within peripheral nerves belong to Schwann cells. Some of the remaining nuclei belong to fibroblasts of the endoneurium, perineurium, and epineurium (i.e., connective tissue) that give tensile strength to the nerve. Perineurium also contains squamous perineural cells (perineural epithelium) which form a continuous layer that isolates the axons within from surrounding connective tissue.
Fibroblast nuclei tend to be smaller and more densely heterochromatic than Schwann cell nuclei, but in most ordinary preparations that include peripheral nerves, it is impractical to distinguish these nuclei.
Note that none of the nuclei visible in peripheral nerves belong to nerve cells. Peripheral nerves do NOT contain nerve cell bodies, only axons of nerve cells whose cell bodies lie elsewhere.
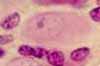
Support cells in peripheral ganglia are sometimes called satellite cells.
Schwann cells can form tumors called schwannomas.
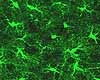
Glial cells -- Support cells of the CENTRAL NERVOUS SYSTEM
The most numerous cells within the central nervous system are glial cells. The name "glia" means "glue" (filling the interstices of nervous tissue), reflecting old but enduring ignorance of their function (and the inadequacy of classical histology to offer much insight). The small nuclei of glial cells may be readily observed in any section of central nervous tissue. Unfortunately, like neurons, these cells are difficult to visualize satisfactorily.
Although glial cells vastly outnumber nerve cells (approx. 10:1, glia:neurons), nerve cells are so large, including the total volume of all their dendrites and axons, that most of the cellular volume of the brain consists of nerve cells.
Ignorance of glial function is beginning to dissipate. For a 2008 review, see:
Barres, BA (2008) The mystery and magic of glia: a perspective on their roles in health and disease. Neuron. 60 (3): 430-40 [PubMedID: 18995817]: This perspective reviews "recent evidence that glial cells are critical participants in every major aspect of brain development, function, and disease. Far more active than once thought, glial cells powerfully control synapse formation, function, and blood flow. They secrete many substances whose roles are not understood, and they are central players in CNS injury and disease. I argue that until the roles of nonneuronal cells are more fully understood and considered, neurobiology as a whole will progress only slowly. ... And please don't forget the glia! Quite possibly the most important roles of glia have yet to be imagined" [emphasis added].
Recent research: In 2025, the journal Science published a report that "astrocytes are indispensible for neuromodulatory signalling across diverse neural circuits, behavioral contexts, and species" (G. Eroglu, "Astrocytes, hidden puppet masters of the brain," Science 388:705-6).
The two most common types of glia, oligodendroglia and astroglia, both have extensive cytoplasmic processes and are intimately involved in the function of nervous tissue. A third glial type, microglia, function similarly to macrophages.
Separately distinguishing among astroglia, oligodendroglia and microglia is a skill for specialists (i.e., pathologists), but with practice their nuclei can be recognized by relative size and texture, with astrocyte nuclei being somewhat larger and paler than the others.
Oligodendroglia (also called "oligodendrocytes" or just "oligos") typically have relatively few processes (hence their name; oligo = few), with each process ending in a sheet of myelin which wraps around a segment of an axon.
Function of oligodendroglia: Oligodendrocytes form myelin in the CNS and hence are responsible for normal propagation of action potentials. Patchy loss of CNS myelin, as in multiple sclerosis, can cause a variety of neurological problems.
Myelin formation by oligodendroglia is slightly different than that by Schwann cells, each of which wraps myelin around a single axon. Each of the several glial cell processes extends to and then myelinates a segment of one axon. If the myelin of one oligodendrocyte process were unrolled, the process would be shaped rather like a wide-bladed shovel (the thin shovel blade would represent the membrane that rolls around the axon to form myelin, and the shovel handle would represent the process which extends back to the glial cell body). Each oligodendroglial cell has several such "shovels," forming myelin around several axons.
Recent evidence from mouse, based on gene transcription profiles, indicates that oligos form several populations. For example, "One population was responsive to motor learning, and another, with a different transcriptome, traveled along blood vessels" (Science, 10 June 2016, 352:1288-1290, DOI: 10.1126/science.352.6291.1288-n).
Astroglia or astrocytes extend branching cytoplasmic processes in all directions (yielding the star-like shape suggested by their name; astro = star). Foot-processes of astrocytes line every surface where central nervous tissue contacts other body tissues, not only the obvious outer surface immediately underlying the pia mater (where they form the glia limitans) but also along every blood vessel and capillary which penetrates into the brain and spinal cord. Other astrocyte foot processes approach nerve cells at any sites where the nerve cell membrane is not otherwise occupied by synapses or by oligodendroglia.
Functions of astroglia: There has been growing awareness this century that astrocytes play several critical roles.
Astrocyte functions and pathologies include all of the following [as of 2003, from Ransom, et al., "New roles for astrocytes (stars at last)," Trends in Neuroscience, 26:520-522, 2003; doi:10.1016/j.tins.2003.08.006].
- Homeostasis, regulating concentrations of K,+ extracellular pH, glutamate and water.
- Maintaining integrity of the blood-brain barrier.
- Modulation of excitatory and inhibitory synapses.
- Neuronal pathfinding during development and regeneration.
- Glioma formation.
- Cytotoxic brain edema.
- Modulation of stroke outcome.
- Hepatic encephalopathy.
- Trophic modulation of neural repair and axon regrowth following injury.
Additional astrocyte functions:
Interactions between central glial cells [astrocytes and microglia] and neurons in the pain circuitry contribute to the pathogenesis of chronic pain [see e.g. Neurotherapeutics (2020) 17:846-860; also see 2021 news article in The New York Times].
Activity of individual astrocytes can correspond closely with that of associated neurons, and can also modulate local blood flow (Schummers, et al., Tuned responses of astrocytes and their influence on hemodynamic signals in the visual cortex, Science 320:1638-1643, 2008; doi:10.1126/science.1156120).
Research has also indicated that astroglia participate in the "glymphatic system" which allows recirculation of CSF and brain interstitial fluid along paravascular channels, a system implicated in sleep (Science news article, 2013).
Serotonin induces changes in gene expression by astrocytes (Science news article, 2023).
"Astrocyte signalling pathways influence neuronal networks and behavioral responses to neuromodulators" (Science perspective, 2025).
Microvascular control: Local variation in blood flow through brain capillaries may be regulated by activity of pericytes, which in turn can respond to neural activity. [Reference: MacVicar & Salter, Neuroscience: Controlled capillaries, Nature 443, 642-643 (12 October 2006) | doi:10.1038/443642a.]
The Blood Brain Barrier
"Blood-brain barrier" is the name given to a physiological property of CNS blood vessels. In contrast to vessels in most other parts of the body, vessels in the brain do NOT allow most molecules to pass freely between blood and interstitial spaces of the brain. The integrity of the blood-brain barrier is established by continuous capillary endothelium together with the absence of endothelial vesicular transcytosis. The only substances which cross this barrier are those which can diffuse through endothelial plasma membranes or those for which specific endothelial membrane channels exist.
The blood-brain barrier is a concept with considerable clinical significance, not only because it limits the delivery of drugs to the central nervous system but also because pathological disturbance of the barrier can seriously impact brain function. For more extensive information, see Blood Brain Barrier at the University of Arizona Health Science Center; at this website, click on ABOUT THE BLOOD BRAIN BARRIER for a drop-down menu.
Microglia are small cells, comprising about 10% of the total brain cell population, which represent the brain's immune system (i.e., macrophage-equivalents residing within the brain). Microglia are also implicated in the maturation, plasticity, and remodelling of synaptic circuits (Science 333:1391, 9 September 2011, doi:10.1126/science.1212112; J. Neurosci. 31 16064-16069, 2011 doi: 10.1523/jneurosci.4158-11.2011)
As described by Kembermann and Neumann (Microglia: the enemy within? Science 302:1689, 5 December 2003, doi:10.1126/science.1092864), the brain exhibits "a robust innate immune response thanks to its microglia, which defend against invading microorganisms and clean up by engulfing the debris of dying cells. In addition, the inflammatory mediators released by microglia during an innate immune response strongly influence neurons and their ability to process information." Recent in vivo observations (Fetler and Amigorena, Brain under surveillance: the microglia patrol, Science 309:392-3, 15 July 2005, doi:10.1126/science.1114852) show microglia as surprisingly dynamic cells, continually extending and withdrawing fine motile cellular processes and contacting astrocytes, neurons, and blood vessels.
Recent research indicates that microglia (in mice) are "an ontogenically distinct population in the mononuclear phagocyte system," originating during embryonic development (Science, 29 October 21, 2010; DOI: 10.1126/science.1194637)
"Interactions between central glial cells [astrocytes and microglia] and neurons in the pain circuitry are critical contributors to the pathogenesis of chronic pain [Neurotherapeutics (2020) 17:846-860]. (Also see 2021 news article in The New York Times.)
Blood vessels in CNS
Central nervous tissue is highly vascular, so blood vessels should be a significant feature in any histological specimen of CNS. Large vessels generally remain on the surface of the brain or spinal cord, so only smaller vessels penetrate into gray and white matter.
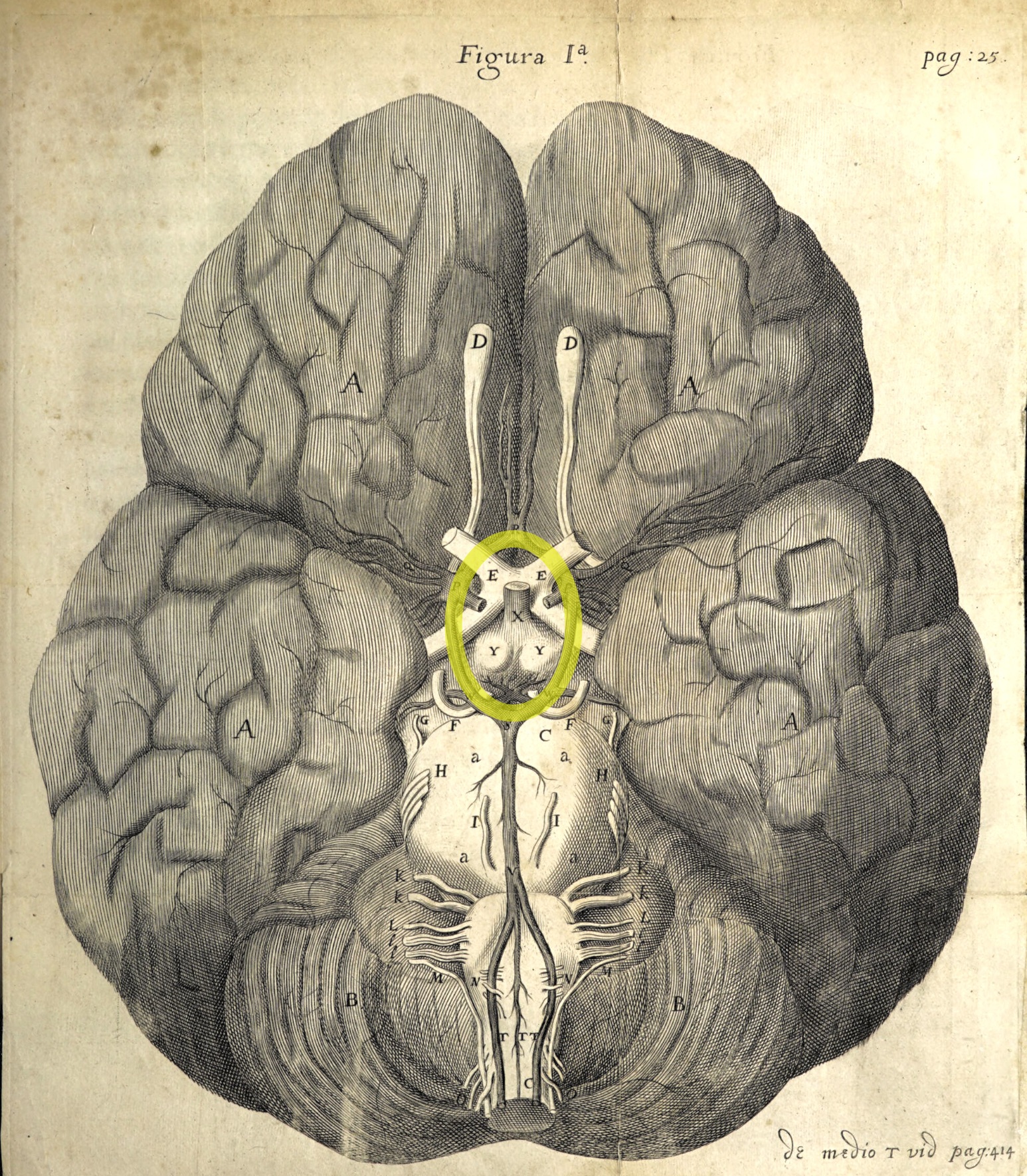
Clinical note: Understanding the course of blood vessels serving various regions of the brain is critical for understanding the effects of stroke when flow through a vessel is compromised.
Historical note: Cerebral vasculature was first described in detail in 1664 in Thomas Willis's Cerebri anatome: cui accessit nervorum descriptio et usus. Willis is commemorated in the circle of Willis, a ring of interconnected arteries at the base of the cerebrum
In histological sections, small vessels may not be immediately recognizable as such. As in other regions of the body, capillaries may be quite inconspicuous due to small size. Even venules and arterioles may be small enough that the layers in their walls are not clearly visible. Blood cells may be washed out during preparation. Nevertheless, such vessels should be noticed, since they play a crucial role in brain function and pathology. (Also see note on microvasculature, above.)
Blood vessels are generally the largest structural elements in both neuropil and white matter (i.e., even capillaries are larger in diameter than most CNS axons and dendrites). The thumbnails below link to several spinal cord specimens in which blood vessels may be observed. Blood vessels appear similar in any region of the brain.
Note that a clear "halo" commonly appears around blood vessels (as well as around cell bodies of neuronal and glial cell bodies). The size of this space is an artifact of histological preparation, resulting from tissue shrinkage when the central nervous tissue is fixed. But it is also a reminder that vessels in the brain are surrounded by usually-inconspicuous perivascular space, which is an extension of subarachnoid space.
 |
 |
 |
 |
|
| Choroid plexus, showing wrinkled epithelium overlying blood vessels, as drawn by A.H. Hassall, 1849, for The Microscopic Anatomy of the Human Body. |
Ependyma, choroid plexus and cerebrospinal fluid
The ventricular system of the brain is lined by a simple cuboidal epithelium called ependyma, a remnant of the embryonic neuroectoderm which once formed the neural tube. At certain sites -- the posterior margin of the lateral ventricles, the midline of the 3rd ventricle, the roof of the 4th ventricle -- this ependyma lies adjacent to overlying connective tissue. Here the ependyma is extensively wrinkled to form choroid plexus, together with blood vessels which are caught up in the folds of ependyma.
Choroid plexus is the source for cerebrospinal fluid (CSF). CSF is actively secreted by the ependymal cells of choroid plexus and (like aqueous humor in the eye) accumulates at a steady rate even if drainage points become occluded.
This is one of three sites associated with the nervous system where a special fluid is produced by a unique tissue, with this fluid needing an outlet elsewhere to avoid buildup of pressure. (The other two sites are the eye and the inner ear. In the eye, aqueous humor secreted by ciliary processes is drained through the canal of Schlemm. In the ear, endolymph secreted by stria vascularis is drained through the endolymphatic sac.) In each of these sites, an imbalance between production and drainage can cause neurological symptoms.
In composition, CSF differs considerably from blood. Although osmolarity and sodium concentrations are similar in blood and CSF, CSF has somewhat more chloride; less potassium, calcium, magnesium and glucose; much less protein, and practically no white blood cells. For specific values as wells as alterations in disease, see Kandel et al., 4th edition, Appendix B, especially pp. 1295-1299.
Research news (2013, 2024): CSF and brain interstitial fluid are exchanged through the so-called "glymphatic system" of paravascular channels. A 2013 report in Science 342:373 implicates this system in the function of sleep (Science news article). A more recent (2024) Science Insight article suggests that "Glymphatic-lymphatic brain cleansing may reveal new therapeutic strategies."
NOTE: This writer is unclear about the distinction between paravascular space and perivascular space. See, for example, here and here.
Research news (2023): Research continues into the immune-related relationships among choroid plexus, CSF, brain interstitial fluid and perivascular channels. For a recent (2023) review (beginning with a short summary), see Science 7 April 2023, p. 52.
The layout of choroid plexus is perhaps most easily appreciated embryologically -- click on the thumbnail for an image of embryonic choroid plexus.
Cerebrospinal fluid accumulates not only from the action of choroid plexus but also from the interstitial spaces of the brain. It flows, under positive pressure developed by its active secretion, through the ventricular system, thence out through holes in the roof of the 4th ventricle into the subarachnoid space, finally draining through "arachnoid villi" into the venous sinuses of the cranial cavity.
Meninges: dura mater, pia mater, and arachnoid
The central nervous system is enveloped by specialized layers of connective tissue.
- The outermost layer is the dura mater (or just "dura"), very dense fibrous connective tissue, tough and fairly impermeable.
- Immediately adjacent to the brain is the pia mater (or just "pia"), a delicate layer of collagen and fibroblast-like cells that adheres closely to the underlying glia limitans (the outermost layer of proper nervous tissue).
- In between dura and pia is the arachnoid, a layer of very loose connective tissue in which cerebrospinal fluid occupies the position of ground substance.
- The name "arachnoid" presumably refers to the spidery, or delicately web-like, network of collagen fibers which extend through the arachnoid layer from dura to pia.
- Perivascular extensions of the subarachnoid space, following blood vessels into the brain, are called "Virchow-Robin space" (the name commemorates Rudolf Virchow, b. 1821, and C.-P. Robin, b. 1821).
- Pia and arachnoid are not distinct, separate layers; together they are sometimes called pia-arachnoid.
- The fluid-filled spaces of the arachnoid layer are sometimes called the subarachnoid space. However, in spite of the "sub" this space is within the arachnoid layer.
- Where the dura envelops cerebral venous sinuses, it is perforated by small passageways called "arachnoid villi" or "arachnoid granulations." These are sites where cerebrospinal fluid drains from the subarachnoid space into venous blood.
SOME EXAMPLES of nervous tissue.
This section offers a guide for microscope lab (i.e., for viewing slides in your reference set). Most of these slides are unlike anything a physician is likely to encounter in practice. What these slides do provide is an opportunity to see for yourself certain features of nervous tissue which are more-or-less readily accessible to microscopic viewing.
Also see:
- Link to virtual slides at LUMEN (Loyola University Medical Education Network). Click on "Part 6: Neural Tissue."
Spinal cord smear
- Each spinal neuron displays classic nerve cell characteristics:
- prominent cell body,
- distinctive round euchromatic nucleus,
- single prominent nucleolus,
- cytoplasm with basophilic masses of Nissl substance (rough endoplasmic reticulum).
The largest nerve cells in this preparation represent spinal motor neurons, the cells whose very long axons extend out peripheral nerves to the muscles. From the nerve cell body extend several dendrites. These are broad at their base and contain Nissl bodies. But dendrites decrease in diameter and basophilia with increasing distance from the soma. The full extent of the dendritic arborization is not visible, since the fine distal branches are obscured in the background texture of the slide.
Each neuron also has a single axon, which can be readily identified only if it begins on the edge of the cell body (as opposed to the top or bottom, as viewed in the slide). The axon, unlike the dendrite, has a uniform diameter and does not contain basophilic Nissl bodies. It begins at the axon hillock, a specialized site on the cell body where the cytoplasm is clear (like the axoplasm, it lacks Nissl bodies). The axon, even more so than the dendrites, disappears into the distance and cannot be followed to its end.
In this same preparation, smaller cells with similar features represent spinal interneurons. Scattered throughout this preparation are also very many cells whose nuclei are smaller than those of the neurons, oval with clumps of heterochromatin, and whose cytoplasm is inconspicuous. These are the glial cells. Numerous capillaries, narrow tubular profiles wandering across the slide, may also be seen.
Spinal cord section
The spinal cord consists of ascending and descending axonal pathways (i.e., white matter) surrounding a central core of gray matter. Use your preferred neuro text to rehearse the functions associated with the following regions in the spinal cord.
- The ventral horns are the broader regions of gray matter which do not extend to the surface of the cord.
- The conspicuous nerve cell bodies in the ventral horn belong to spinal motor neurons. These spinal motor neurons are lost in ALS, amyotrophic lateral sclerosis.
- The relative size of the ventral horn in any particular section depends on the level in the spinal cord where the section was taken. The ventral horn is bulkier at levels that connect with arms or legs.
- Dorsal columns are the white-matter fiber tracts between the dorsal horns.
- Lateral and anterior columns are the white-matter fiber tracts which form the sides and front of the cord.
- White matter columns are thicker (containing more axons) in the upper cord, reflecting the number of axons at each level. (For example, as one ascends the cord, more and more sensory axons join the dorsal column.)
- The central canal is the small channel within the "bridge" of gray matter which connects the gray matter of left and right sides. The central canal should properly be lined by ependyma (epithelial tissue), but this is seldom visible on our slides.
Some sections of spinal cord may include dorsal and ventral roots containing (respectively) sensory and motor axons.
- Sensory axons in a sensory (dorsal) root enter the cord at the dorsal horn. Cell bodies of sensory axons occur in dorsal root ganglia, located near the cord along the dorsal root.
- Motor axons in a motor (ventral) root originate from motoneuron cell bodies in the ventral horn and pass through white matter before leaving the cord.
Cerebral cortex
The cerebral cortex forms the surface of gyri and sulci over each entire cerebral hemisphere. Its composition is complex (after all, it is the seat of conscious perception and thought!), with many different types of nerve cells. These include many local interneurons (stellate cells and granule cells) as well as the much larger and more conspicuous pyramidal cells, some of whose axons enter the underlying white matter and travel to other cortical areas or to other regions of the brain.

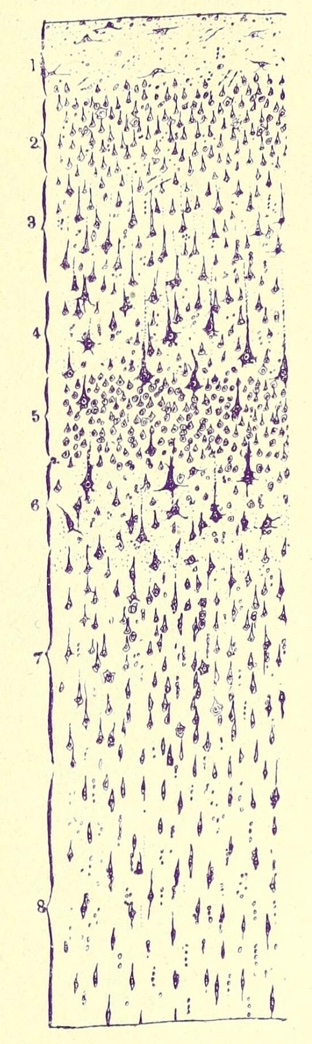
The cerebral cortex is traditionally (but rather arbitrarily) described as having six layers. Functional localization in the cortex correlates with noticeable differences in the numbers and sizes of cell bodies in these several layers, now mapped as Brodmann's areas. Although the several layers are not clearly demarcated (they are arbitrary, after all), they can be roughly approximated by looking for the following features.
Layer I (the "molecular layer") is the outermost layer. This layer contains relatively few nerve cell bodies. The odd name "molecular layer" derives from the fine texture of this layer, due to its composition largely of dendrites and fine axon terminals (and glia, of course). This layer might possibly have a special role in memory (Science 374:538, 20 Oct. 2021).
Layer II (the "outer granular layer"), typically contains many very small cells (granule cells).
Layer III (the "outer pyramidal layer") contains cell bodies of small pyramidal cells. Axons from these cells typically project to the upper layers of neighboring cortical regions.
Layer IV (the "inner granular layer") contains axonal ramifications of afferent fibers, such as sensory axons from the thalamus. Axons from the lateral geniculate nucleus (the visual relay of the thalamus) are so numerous that the primary visual cortex which receives these axons (Brodmann's area 17, at the occipital pole of each hemisphere) is sometimes called "striate cortex," because these axons conspicuously divide the cortex into layers that are visible to gross inspection.
Layer V (the "inner pyramidal layer") contains cell bodies of large pyramidal cells. Axons from these cells typically project to more distant cortical regions, to other parts of the brain, or to lower centers (such as spinal motor neurons). The larger size of these pyramidal cell bodies (compared the the smaller cells of layer III) is associated with the greater length of their axons. (Recall that cell bodies provide most of the basic cellular functions needed to maintain the axon, while the axonal surface membrane and axoplasmic volume may be many times greater than the surface and volume of the cell body.)
Layer VI (the "layer of pleiomorphic cells) typically contains cells of assorted size and shape (hence, "pleiomorphic").
Deep to layer six is white matter containing axons going to and from the cortex.
Historical note: Regional variations across the cortex in the "cytoarchitecture" (detailed histological appearance) of these several cortical layers were described over a century ago by Korbinian Brodmann (b. 1868). Brodmann's descriptions formed the original basis for recognizing Brodmann's areas, now known to correspond with functional localization in the cortex.
Cerebellar cortex
The cortex of the cerebellum consists of three well-defined layers. The most prominent nerve cells are Purkinje cells, whose cell bodies all lie at the same level.
The outer molecular layer consists principally of the dendrites of Purkinje cells and the axons of granule cells. The odd name "molecular layer" derives from the fine texture of this layer, due to its composition largely of dendrites and fine axon terminals. Nuclei in this layer belong mostly to glial cells.
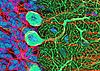
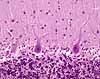
The Purkinje cell layer contains large cell bodies of Purkinje cells, the sole output cells for the cortex.
The inner layer, or granule cell layer, is packed with nuclei of vastly many cerebellar granule cells. These are among the smallest (and most numerous) neurons in the body.
Deep to the granular cell layer is white matter containing axons going to and from the cortex.
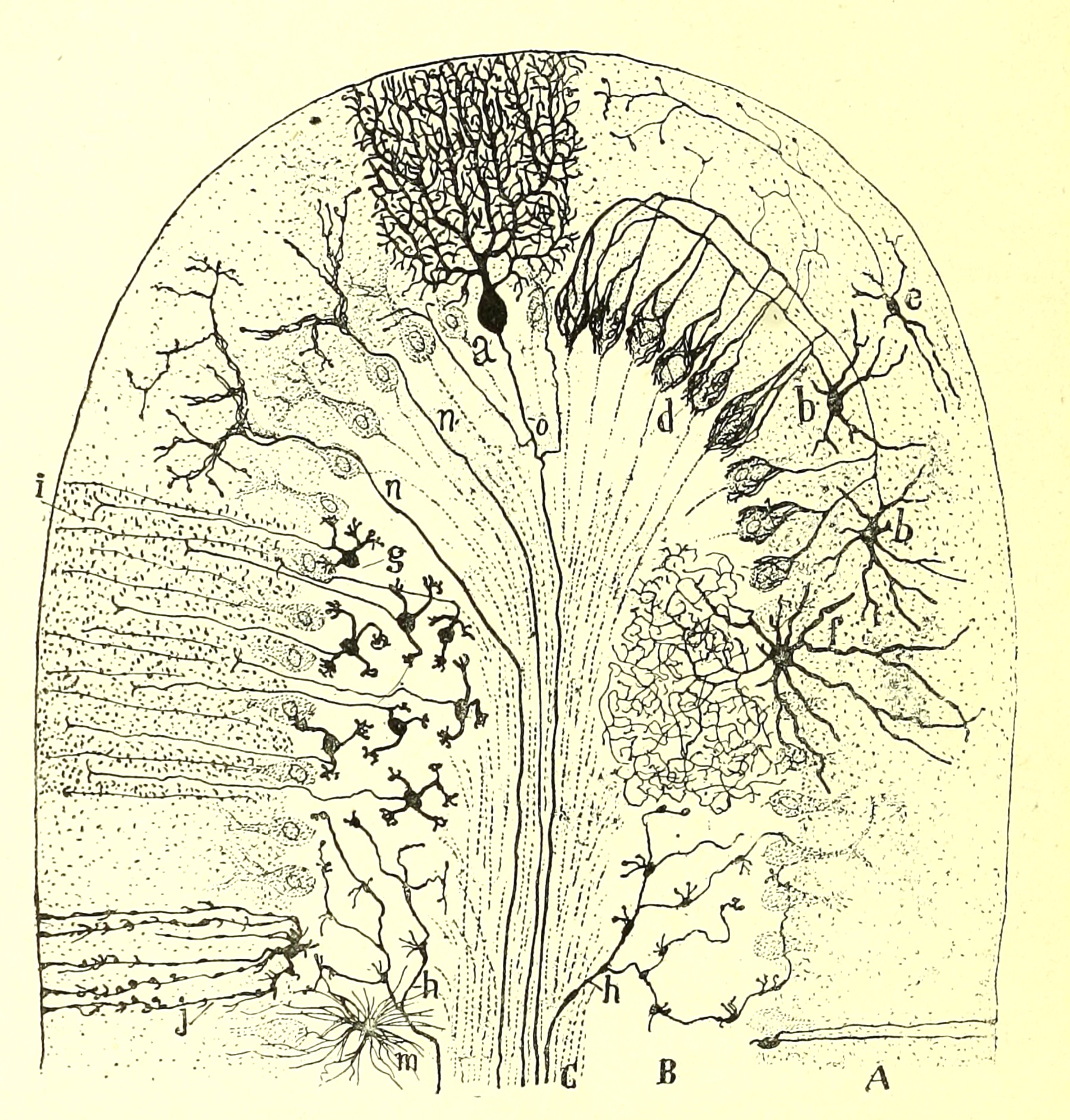
The pattern of connections among various axons and dendrites in the cerebellum is extremely elegant and regular, and has been described in extensive detail. Any thorough neuro text (e.g., Kandel et al., 4th ed., pp. 835 ff) should have a good account.
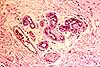
Peripheral ganglia
Both the paravertebral ganglia of the sympathetic nervous system and the scattered ganglia of the parasympathetic nervous system consist of small clusters of nerve cell bodies.
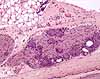
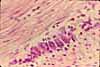
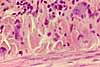
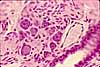
Paravertebral dorsal root ganglia contain the cell bodies for somatic sensory nerves.
Parasympathetic ganglia may turn up in sections of various visceral organs, where they can be recognized by the classic appearance of nerve cell bodies.
Retina
Tissues of the eye are listed in a separate page.
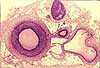
Composition and appearance of PERIPHERAL NERVES
Like other portions of the nervous system, peripheral nerves are a part of a functioning, highly organized whole. Each portion must be understood in relation to the rest of the system.
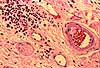
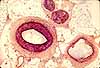
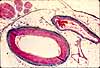
Examples of peripheral nerves are often fairly easy to find in sections of the skin. Larger nerves often run in parallel with blood vessels.
Click on a thumbnail below for enlargement.Peripheral nerves consist of axons bundled together within an epineurium, or connective tissue sheath. The collagen here confers ess ential tensile strength to the nerve, since axons themselves are quite delicate.
Clinical note: The epineurium gives surgeons something to hold onto and suture through when reattaching severed nerve ends.Peripheral nerves are functionally meaningful only in relation to their connections. All of the axons which travel along peripheral nerves begin and end somewhere else.
Motor axons originate with cell bodies in the spinal cord's ventral horn or in the brainstem's motor nuclei or in peripheral sympathetic or parasympathetic ganglia. Motor axons terminate at muscles (including smooth muscle along blood vessels) or glands.
Somatosensory axons begin with a peripheral receptor (e.g., a Meissner's corpuscle in skin or a muscle-spindle in a muscle). These sensory axons then travel toward their cell bodies in a dorsal root ganglion or trigeminal ganglion, and finally terminate at synapses within the spinal cord or brain stem. (Note that somatosensory axons are an exception to the rule that axons always conduct impulses away from the cell body.)
All the cellular nuclei which are obviously visible within a peripheral nerve belong not to nerve cells but to Schwann cells or to fibroblasts.
In routine H&E slides, three types of fibrous tissue bear some resemblance to one another: peripheral nerves, smooth muscle, and collagen. All three are eosinophilic, and all three contain scattered, elongated nuclei.
Several features may be used to distinguish nerves from smooth muscle or other fibrous tissue.
- Nerves have a sheath of fibrous connective tissue, the epineurium, that forms a discrete boundary around the nerve. Small nerves, and bundles of axons within large nerves, are also ensheathed by perineurium, including squamous cells (perineural epithelium) whose intercellular junctions isolate the axons from surrounding connective tissue.
Clinical note: The epineurium gives surgeons something to hold onto and suture through when reattaching severed nerve ends.- Unless specially stained, nerve fibers tend to appear rather pale.
- Nerves often have a characteristic swirled or wavy texture, because axons in the nerve tend to twist, like fibers in a string.
- Axons need to be somewhat longer than the nerve within which they run, so that if the nerve is stretched the axons do not all snap.
- The nuclei found within nerves are mostly Schwann cell nuclei.
- Schwann cell nuclei are usually larger and with less-condensed chromatin than fibroblast nuclei.
- A small bundle of smooth muscle can resemble a nerve, but smooth muscle bundles have no sheath. With commonly used stains, smooth muscle is typically colored more intensely than nerve.
- In longitudinal section, smooth muscle nuclei typically appear considerably longer than either Schwann or fibroblast nuclei, and in cross section of well-prepared specimens these nuclei can usually be seen to reside within the fibers (unlike either Schwann cell or fibroblast nuclei, which lie alongside the associated fibers).
Note that the texture of peripheral nerves can differ from site to site, depending on axon size and especially on the proportion of myelinated to unmyelinated axons. For example, nerves in the tongue, with many large myelinated axons, are much more obvious than are autonomic nerves in Auerbach's plexus of the gut, where most axons are smaller and unmyelinated.
In ordinary H&E stained cross sections of peripheral nerve, myelin might be visible as a pale unstained halo around a larger axon. Less-than-ideal fixation often distorts the relationship, so the axon may not be centered within the halo and the myelin itself easily mistaken for extracellular space.
Many details of peripheral nerves cannot be well-appreciated by light microscopy. For electron micrographs of peripheral nerves, see the online Electron Microscopic Atlas of cells, tissues, and organs (the text is in German, but most figure labels can be deciphered fairly easily).
SENSORY and MOTOR NERVE ENDINGS associated with PERIPHERAL NERVES
For sensory receptors in skin, see skin innervation. For sensory receptors associated with muscle, see muscle innervation. Cell bodies for these and most other somatosensory endings are located in dorsal root ganglia.
For motor endings on skeletal muscle, see muscle innervation. Cell bodies for these motor endings are the spinal motor neurons in the anterior horn of the spinal cord.
Composition of CENTRAL NERVOUS SYSTEM
The organization of the central nervous system is based upon interconnections across varying distances among billions of individual nerve cells. The basic principle of neural organization is quite straightforward: Nervous tissue consists of nerve cells communicating with other nerve cells. This simple yet fundamental concept can easily become lost in the forest of details presented in standard textbooks. Here, then, is a brief guide to nervous tissue, including the classification and nomenclature of nerve cells.
Gray Matter and White Matter (Cortex, Nuclei, and Fiber Tracts)
- Regions consisting of cell bodies together with their associated dendrites and axon terminals are termed gray matter.
- Gray matter located on the surface of the brain is called cortex (e.g., cortex of the cerebral hemispheres, cortex of the cerebellum).
- Masses of gray matter located deeper in the brain are called nuclei.(e.g., brainstem nuclei, nuclei of the thalamus).
- Regions consisting of axons gathered into bundles, to the exclusion of cell bodies, are called white matter.
- White matter in which all of the axons lie parallel to one another is called a fiber tract.
- A fiber tract which crosses the midline to connect bilaterally symmetric structures is called a commissure.
The microscopic appearances of gray matter and white matter may be conveniently contrasted in a section of spinal cord.
Various stains have variously differential effects on gray and white matter. Note that a popular neuroanatomical stain (Weigert's stain, commemorating Karl Weigert, b. 1845), used to highlight different brain regions, colors myelin black. Thus, paradoxically, in many pictures of the brain, white matter appekars black while gray matter appears pale.
- Living gray matter is not gray but rather pink, due to blood perfusing through very numerous brain capillaries. (The brain is intensely vascular, with each cubic centimeter of brain tissue having around 100 square centimeters of capillary endothelial surface area.)
A literary aside: "Gray matter" is colloquially used as an expression for "smarts." But, Hercule Poirot notwithstanding, there is no such thing as "little gray cells."- Cortex is gray matter found on the surface of the brain. There is cerebral cortex covering the surface of cerebrum and cerebellar cortex covering the surface of the cerebellum. But not all gray matter is cortical.
- A nucleus is a mass of gray matter found deep in the brain. Such nuclei are not to be confused with nuclei of individual cells, although neuron cell bodies with their cell-nuclei are found within brain-nuclei (and not in white matter).
Note that gray matter necessarily contains both the beginnings and endings of axons, even though the greater portion of many axons' length is contained within the fiber tracts of white matter. Gray matter is gray not because it lacks myelin, but because it contains lots of other stuff besides myelinated axons.
White matter: Axons from many different neurons often gather together in large numbers at some distance from their cell bodies. In such regions, the relatively large amount of myelin confers a white color, hence, white matter. Myelin is largely fat, which is white in both living and fixed condition.
NOTE: In many neuroanatomical images, white matter has been stained black.
Clinical note: White matter is selectively involved in some disorders. See the journal Science 372:6548 (18 June 2021) for an essay on white matter per se.
- White matter represents axons going relatively long distances. It is the stuff of "fiber tracts" or "neural pathways."
- If white matter is cut, the cell body at one end of each axon is disconnected from its distal axon terminals at the other end.
- In some white matter areas most axons are travelling in parallel, with all action potentials propagating in the same direction. For example, most axons in the dorsal columns are ascending, while those in the cortico-spinal tract are descending.
- But in many other white matter regions of the CNS, adjacent axons may carry signals in opposite directions or be interwoven in a complex meshwork. For example, axons in the internal capsule and corpus callosum crisscross back and forth, interconnecting many different regions of the cerebral hemispheres.
Although white matter consists of myelinated axons (and unmyelinated axons as well), myelinated axons are not excluded from gray matter. Myelinated axons must begin and end somewhere, and that place is with cell bodies and dendrites of gray matter. Gray matter just has a lot of other stuff in it besides myelinated axons.
Sensory Neurons, Motor Neurons, and Interneurons
Sensory neurons convey sensory information into the central nervous system. Primary sensory neurons receive their information directly through sense receptors rather than dendrites. Second-, third- and higher-order sensory neurons relay information to sequentially higher levels in the brain.
Primary sensory neurons have their cell bodies located outside the central nervous system. For example, most somatosensory cell bodies reside in dorsal root ganglia (or in the trigeminal ganglion).Motor neurons (or motoneurons) convey information out from the central nervous system to muscles or glands. Lower motor neurons, located in the ventral horn of the spinal cord or in motor nuclei of the brainstem, send their motor axons out peripheral nerves. Upper motor neurons, pyramidal cells located in the motor cortex, relay information to the lower motor neurons.
All other neurons are interneurons. They interconnect neurons with other neurons. Nearly all the nerve cells in the central nervous system are interneurons. Their axons arise in one region of the CNS where the cell body resides and end somewhere else (sometimes several other places). Second order, third order, and higher order sensory neurons can be considered as ascending interneurons; upper motor neurons can be considered as descending interneurons.
Synaptic Relays
Information from primary sensory neurons does not reach the highest levels (the cerebral cortex) directly. Rather it is relayed at least twice (once in the spinal cord or brain stem, again in the thalamus). At each relay, incoming (afferent, presynaptic) axons terminate by synapsing onto the dendrites of the next neurons in the series. The outgoing axons of these neurons then relay the information to the next level. At each relay site, some information processing and distribution can occur, so the information can be altered as it travels upward. Similarly, muscle commands are relayed downward from motor cortex and other motor centers to the "final common pathway," the lower motor neurons of cranial nerve nuclei and the anterior horn of the spinal cord.
Because each relay occurs at synapses onto dendrites and cell bodies of the next neurons in the pathway, each relay is associated with gray matter. Conversely, every gray matter region (nucleus or cortex) is associated with relaying information from one set of axons (the afferent axons that enter the region in question) to another (the efferent axons that leave the region).
Sometimes it is sufficient just to know the beginning and ending points of an entire pathway. Other times knowing how far the neurons of each relay extend will be necessary to determine the site or effects of a lesion.
Afferent and Efferent Axons
All gray matter regions of the brain, both cortex and nuclei, are associated with afferent ("input") and efferent ("output") axons. Afferent axons enter the region from somewhere else (i.e., the cell body is located some distance away). Efferent axons arise from cell bodies within the region and leave the region to go somewhere else. Thus every long-distance axon is both efferent (with respect to its source, the location of its cell body) and afferent (with respect to its destination).
The terms "afferent" and "efferent" are relational terms. Neither can be used precisely without specifying a region of reference. For example, "cortical afferents" provide input to cortex; they may be efferents from thalamus or efferents from some other cortical area.
Ascending and Descending Pathways
"Ascending" and "descending" refer to directions along the neural axis. These terms may often correlate with "afferent" and "efferent," at least when the reference is high, like cortex. (In fact, "afferent" and "efferent" are sometimes used as synonyms of "ascending" and "descending," respectively. But they also have a relational meaning, defined above. "Ascending" and "descending" are also closely associated with "sensory" and "motor," respectively. But both sensory and motor information can be passed up, down, and sidewise, so these words should not be carelessly interchanged.
Long-Axon and Short-Axon Neurons
Gray matter typically contains both many short-axon neurons and a smaller number of long-axon neurons.
Long-axon neurons (also called principal cells, projection neurons, or Golgi type 1 neurons) have the largest and most conspicuous cell bodies in a particular region of gray matter. These cells generally have very long axons which leave the local region to project elsewhere, usually traveling within some white matter fiber tract. These long axons may extend for appreciable distances, up to many centimeters. Long-axon neurons are responsible for communicating with other brain regions. Every parcel of gray matter has a class of long-axon neurons; otherwise information would come in but never go out. The axons of a region's long-axon neurons are by definition identical with the region's efferent axons.
- Such large, long-axon cells may have specific names like pyramidal cells (the output cells of the cerebral cortex) or Purkinje cells (the output cells of the cerebellar cortex).
- A large cell body is probably necessary to support the cytoplasmic volume and membrane surface area of an extremely long axon. (For exercise, you might try estimating the volume of axoplasm in an axon 5 �m in diameter and ten centimeters long. Compare that result with the volume of cytoplasm in a cell body 50 �m in diameter.)
The study of neuroanatomy is largely the study of the axonal projections of long-axon cells.
Historical note: See "The study of neuroanatomy" above.Short-axon neurons (also called intrinsic neurons, local interneurons, or Golgi type 2 neurons) have relatively short axons that do not leave the immediate neighborhood.
- Short-axon neurons are presumably responsible for integrating information from diverse sources. ("Integrating" is the jargon term for what nervous tissue does when it transforms information, or thinks. The word "integration" provides a convenient name for a process that is otherwise poorly understood.)
- Cell bodies of short-axon neurons are often small and numerous. They come in various shapes, and are given names according to the region where they are found (see neuron names, below).
- Most so-called granule cells and stellate cells are local interneurons.
In most regions, long-axon cells are much better understood than intrinsic cells. Long axons provide opportunity for researchers to record and interfere with neuronal output. Cells with short axons are much more difficult to manipulate.
Historical note: The terms "Golgi type 1" and "Golgi type 2" commemorate pioneering neuroscientist Camillo Golgi (b. 1843).
Nerve cell types and nerve cell names
Unlike many other cell types in the body (e.g., epidermal keratinocytes or skeletal osteocytes), nerve cells are not all equivalent to one another. Neurons in one region are structurally and functionally different from those in other regions, with different sources of input, different destinations for output, different patterns of dendritic branching, different neurotransmitters, etc. Similar diversity is found even within a given region (e.g., cerebral cortex and cerebellar cortex, above). The result is a tremendous variety, with specific names for quite a few distinctive nerve cell types.
Indeed, in many animals certain neurons can each be identified as a unique individual cell, with features that distinguish that one particular cell from any other cell in the same animal. Such identifiable neurons can be consistently recognized from specimen to specimen, sometimes even in specimens from related species.
Historical note: The great variety of nerve cells was famously observed and described by Ramón y Cajal from the late 1800s through the early 1900s. "So varied were the cells' structures in the human cortex -- some look like stars, others like bird's nests, baskets, or spindles -- that Cajal divided them into dozens of subtypes. He argued that the diversity of the cells, which he called the 'butterflies of the soul,' was the key to higher cognition" (quote from Science 349:575, 7 Aug 2015).
Recent research: In 2021, the journal Nature 598:66 (6 October 2021) provided an initial progress report on "a multimodal cell census and atlas of the mammalian primary motor cortex as the initial product of the BRAIN Initiative Cell Census Network." In this report, "cells grouped according to their transcriptomes tend to share other features, such as location, shape, and electrical activity. That finding 'provides strong validation to the molecularly defined cell types... For the human motor cortex, 127 such types emerged'" (quote from Science daily news, 7 October 2021). Note that those 127 cell types were all from one single -- and rather small -- brain region, the primary motor cortex.
This work on the BRAIN Initiative Cell Census continues apace, with multiple articles published on Oct. 13, 2023, in a special issue of Science. For a nice news media report by Carl Zimmer, see The New York Times, 12 Oct. 2023: "Researchers identified some 3,300 types of brain cells, an order of magnitude more than was previously known, and have only a dim notion of what most of them do."
Nerve cells in the brain of Drosophila have been much more thoroughly mapped [Nature 634: 139-152, 2024; New York Times article by Carl Zimmer, October 2, 2024], with 140,000 neurons of 8,453 cell types.Proper synaptic connections between specific neurons is guided by a "barcode" of cell-surface proteins; see Science 385:370 (26 July 2024).
As a result of this diversity of nerve cell types, neurohistology is burdened by a profusion of nerve cell names. Every nerve cell can be classified according to its place within the general organization of nervous tissue (above). But each nerve cell also belongs to a unique population with a particular role in the information processing of the brain.
Examples of a few nerve cell types, familiar from many textbook accounts, are described below.
Neuron preparation by Ramón y Cajal
Pyramidal cells are the efferent (long-axon) cells of the cerebral cortex. The name refers to the shape of the cell body as seen in standard sections perpendicular to the cortical surface. The apex of the pyramid points toward the cortical surface. A large apical dendrite extends farther upward toward that surface, while other dendrites arise from the corners and sides of the pyramid. The axon extends down into white matter (the internal capsule) from the base of the pyramid. Most pyramidal cells project association fibers to other cortical regions and/or to deeper nuclei of the brain. (Classic illustrations of these cells were done by Ramón y Cajal).
Stellate cells are intrinsic neurons named for their star-like shape, which results from dendrites arising in many directions. The name is descriptive but relatively non-specific. Thus stellate cells of the cerebral cortex are not the same as stellate cells of the cerebellar cortex.
Horizontal cells are intrinsic neurons whose dendrites and local axons tend to be confined within a layer parallel to a surface. Again the name is descriptive rather than specific. Thus horizontal cells in the cerebral cortex are not the same as horizontal cells in the retina.
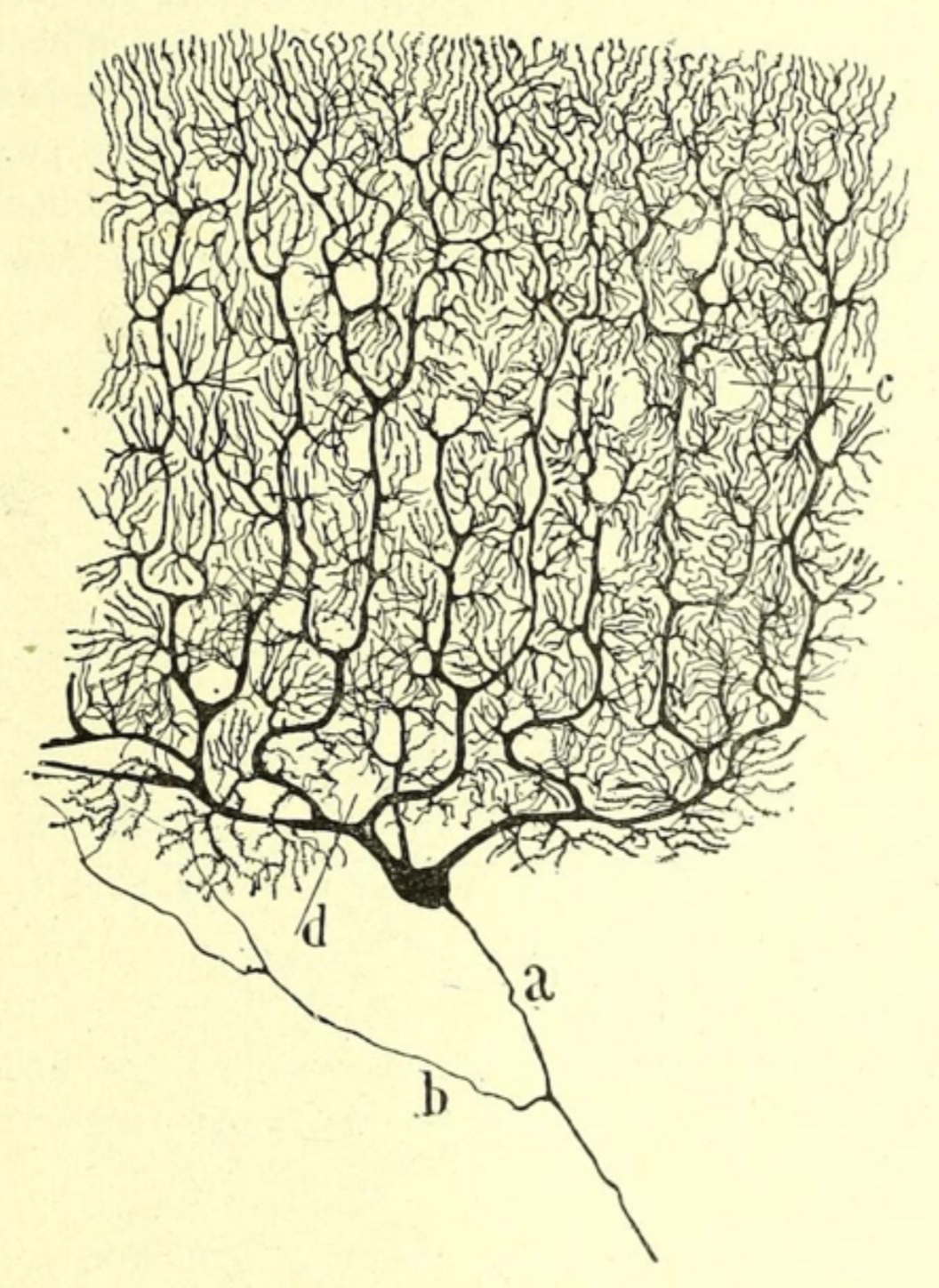
The synaptic input of Purkinje cells comes largely from the cerebellar parallel fibers (which are the axons of the extremely numerous local interneurons, the cerebellar granule cells). Purkinje cells also receive potent synapses directly from one class of cerebellar afferent fibers, the "climbing fibers." (Another class of cerebellar afferents, the "mossy fibers," provide input to the intrinsic granule cells. In addition to Purkinje cells and granule cells, the cerebellar cortex also contains stellate cells, basket cells, and Golgi cells.)
[See your favorite neuro text (e.g., Kandel et al., 4th ed., pp. 835 ff) for elegant details of neural circuitry in the cerebellar cortex.]
Spinal motor neurons have the largest nerve cell bodies in the ventral horn. These cells are also called "lower motor neurons," or just "motor neurons" (since upper moter neurons are properly called interneurons). Axons of these cells extend through ventral (anterior) roots into peripheral nerves, and hence to motor end plates on muscle fibers. Each muscle fiber of skeletal muscle is innervated by a single spinal motor neuron. Each spinal motor neuron innervates one or more muscle fibers; the number of muscle fibers per axon is related to the fineness of motor control -- e.g., one-to-one for eye muscles, many-to-one for large postural muscles. A single spinal motor neuron and all the muscle fibers innervated by its axon are called a motor unit. (In the cartoon diagram to right, a spinal motor neuron is represented extending into the figure's right leg to innervate a calf muscle. The upper neuron in this cartoon, whose cell body is located in the head, represents an upper motor neuron, or pyramidal cell, of motor cortex.)
Dorsal root ganglion neurons (which are primary somatosensory neurons) are among the most distinctively shaped of our nerve cells. Their cell bodies are peculiarly located in dorsal root ganglia alongside the spinal cord. A single process leaves each cell body, then bifurcates into two very long branches; both branches together comprise a single functional axon. The distal branch serves one of many peripheral touch or proprioceptor sensory nerve endings. The proximal branch enters the spinal cord and contributes to one of the ascending afferent tracts, with some interneuronal connections along the way. (In the cartoon diagram to right, a dorsal root ganglion neuron is represented extending from the figure's left toe up to the figure's neck.)The above are only examples, from classical textbook literature. Every region of the central nervous system contains many distinct neuronal cell types, most of which are appreciated only by the research specialist. But a few such examples are worth knowing, if only because they are a part of familiar scientific knowledge and because they hint at the complexity of neural organization.
Note that in every region of the brain, the long-axon cells are directly associated with significant neuroanatomical pathways (output), while detailed knowledge of the intrinsic cells generally has little clinical importance -- at least in our current state of ignorance.
NERVOUS TISSUE PATHOLOGY.
Washington University hosts an excellent web resource for neuromuscular pathology.
Within this site, you can find Differential Diagnosis for myopathy and neuropathy, and much more.Chapter 18 in the 3rd edition of Kandel et al. (1991) is an excellent resource for understanding the responses of nervous tissue to injury. The newer, 4th edition of Kandel et al. (2000) offers a much sketchier account in Chapter 55 (pp. 1108 ff).
WebPath also offers some examples of nervous system pathology, see WebPath CNS Pathology Index and WebPath CNS Degenerative Diseases.
For example:
- Spinal motor neurons are lost in amyotrophic lateral sclerosis (WebPath, gross; WebPath, neuron loss; WebPath, gliosis).
- Alzheimer's disease (WebPath) is accompanied by cortical changes (WebPath gross, WebPath, WebPath, WebPath, WebPath, WebPath, WebPath). Each of these links offers a different illustration.
- Inflammation, as in meningitis (WebPath ), presents infiltration of leukocytes into regions, such as CSF, where they are normally absent.
- Multiple sclerosis (WebPath, WebPath w/ MRI ) involves patchy loss of CNS myelin, apparently caused either by autoimmune destruction of oligodendroglia or by malfunction of oligodendroglia with immune destruction as a secondary effect (Science 308:778, 2005). Resulting loss of conduction can cause a variety of neurological problems.
- "Microscopically, the caudate nucleus in Huntington's disease demonstrates loss of neurons along with gliosis" (WebPath, gross; WebPath, microscopic).
- Parkinson's disease (WebPath ) involves loss of the pigmented neurons in the brainstem which give substantia nigra ("black substance") its distinctive appearance.
- Brain tissue in CJD or Creutzfeldt-Jakob disease (WebPath, WebPath, WebPath) is characterized by microscopic vacuoles (hence, "spongiform encephalopathy") and plaques.
- Schwann cells can form tumors called schwannomas (see WebPath: MRI, gross, dissection, microscopic low X, high X).
Comments and questions: dgking@siu.edu
SIUC / School
of Medicine / Anatomy / David
King
https://histology.siu.edu/ssb/neuron.htm
Last updated: 9 September 2025 / dgk