
Liver
INTRODUCTION TO LIVER
The liver is one of the largest, most important, and least appreciated organs in the body. The bulk of the liver consists of hepatocytes, which are epithelial cells with a unique shape and a unique configuration.
The liver is essentially an exocrine gland, secreting bile into the intestine. But the liver is also -- and significantly so -- an endocrine gland and a blood filter. The liver has a diversity of functions not typically associated with glands. The liver is a metabolic factory, synthesizing and breaking down a variety of substances. It's functions include all of the following:
- formation and secretion of bile.
- storage of glycogen, buffer for blood glucose.
- synthesis of urea.
- metabolism of cholesterol and fat.
- synthesis and endocrine secretion of many plasma proteins, including clotting factors.
- detoxification of many drugs and other poisons.
- cleansing of bacteria from blood.
- processing of several steroid hormones and vitamin D.
- volume reservoir for blood.
- catabolism of hemoglobin from worn-out red blood cells.
Much of the liver's histological organization is designed to support the liver's central role as a blood filter.
Housecleaning
An analogy for liver and kidney function.The body contains two "blood-filter" organs, the liver and the kidney. They both serve to remove unwanted materials from blood, but they do so by two very different methods. Without going into any molecular detail, we might compare these two methods to the strategies adopted by two different householders who set out to eliminate clutter from their respective homes.
One householder identifies each unwanted item and tosses it into the trash. Only materials identifiable as trash are disposed of. This householder works like the liver, where hepatocytes take up various molecules from the blood and destroy or detoxify them as needed.
The other householder first puts everything out into the yard, then identifies and retrieves anything that was worth keeping. Only materials identifiable as valuable are retrieved. This householder works like the kidney, which lets practically everything pass out from blood into glomerular filtrate and then uses proximal tubules to actively pump any valuable molecules back into renal capillaries.
The liver receives a dual vascular supply.
- The hepatic portal vein brings to the liver all of the blood which has previously passed through the intestine and spleen.
- The hepatic artery brings fresh, oxygenated blood from the aorta.
Portal venous blood from the intestine and spleen and arterial blood from the aorta mix together in hepatic sinusoids before leaving the liver in the hepatic vein.
The liver receives over 25% of the total resting cardiac output and is responsible for over 20% of the body's resting oxygen consumption.
The liver is organized into lobules which take the shape of polygonal prisms. Each lobule is typically hexagonal in cross section and is centered on a branch of the hepatic vein (called, logically enough, the central vein).
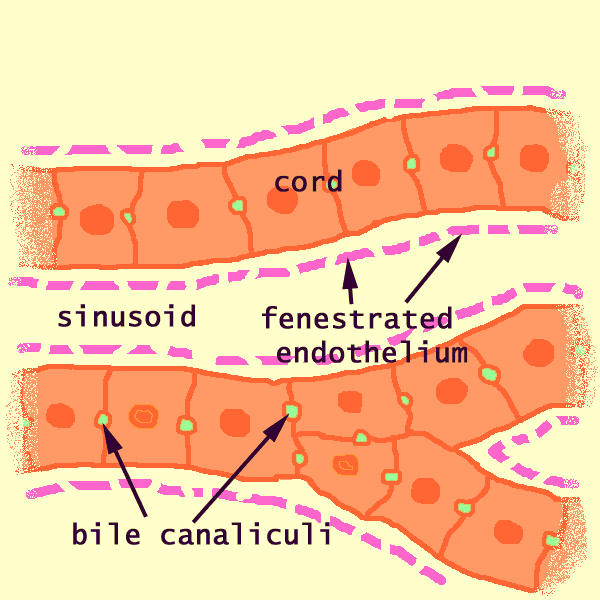
Within each lobule, hepatocytes are arranged into hepatic cords separated by adjacent sinusoids.
The fenestrated endothelium lining the sinusoids lies immediately adjacent to the cords, with no basement membrane and practically no intervening connective tissue, so that each hepatocyte is bathed on two faces by blood plasma.
Although the liver is an exocrine gland (with bile duct leading to the intestine), the cells of the liver are NOT organized into secretory acini or tubules. Instead, hepatocytes form linear cords, within which a network of bile canaliculi provides passage through intercellular channels to the nearest branches of the bile duct.
This remarkable tissue arrangement appears to optimize the liver's several roles as exocrine gland, endocrine gland, and blood filter.
Dual vascular supply.
All of the blood which passes through the intestine and spleen is delivered to the liver by the hepatic portal vein.
This portal blood carries not only nutrients but also various contaminants (drugs, toxins from food, bacteria, byproducts of blood-cell recycling) which have been absorbed through the intestinal mucosa or produced in the spleen.
The liver also receives arterial blood, carrying oxygen, from the hepatic artery.
Blood from both portal vein and hepatic artery mixes together in the hepatic sinusoids and then passes out of the liver through the hepatic vein. The parenchyma of each lobule can be divided into arbitrary zones based on oxygen supply (see WebPath), with the central zone (closest the the central vein) poorest in oxygen.
This blood flow pattern can yield visible differences in hepatocyte appearance. For example, glycogen is depleted first from the periphery during times of fasting and deposited first in the periphery during times of feasting. This leads to a pattern of glycogen which can reflect recent nutritional history.
Both the hepatic portal vein and the hepatic artery branch in parallel along the corners of hepatic lobules, in regions called portal areas.
A branch of the hepatic vein, called the central vein, runs along the central axis of each lobule.
Failure of adequate venous drainage leads to chronic passive congestion.
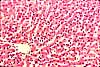
Organization of Liver Lobules
The liver is organized into lobules (portal lobules, hepatic lobules) which take the shape of irregular polygonal prisms. Some texts refer instead to liver acini; see below.
At the corners between adjacent lobules are the so-called portal areas (portal canals, portal triads). These are regions of connective tissue which include branches of the bile duct, the portal vein, and the hepatic artery.
Along the central axis of each lobule runs a central vein, which is a branch of the hepatic vein.
Occupying the bulk of the lobule are hepatocytes arranged into cords. separated by sinusoids.
Lobules appear quite clearly in pig liver, which has an envelope of fibrous connective tissue around each lobule. (This tough connective tissue is one reason why pig liver, unlike calf liver or chicken liver, is not a popular menu item.)
The lobular organization of the human liver is not immediately evident under the microscope. Lobules do not have distinct boundaries, and they are seldom cut neatly in cross section.
Increased quantity of portal connective tissue is indicative of cirrhosis. Increased numbers of leukocytes in portal areas is indicative of hepatitis.
Failure of adequate venous drainage leads to chronic passive congestion.
Some texts refer to liver acini rather than liver lobules. These are both arbitrary ways to conceptualize the same tissue.
- As described above, a liver lobule encompasses the liver tissue that is served by single branch of a central vein (which is a branch of the hepatic vein). The lobule is roughly hexagonal in cross section, with a central vein at its center and portal areas at its peripheral corners.
- In contrast, a liver acinus encompasses the liver tissue that is served by a single terminal branch of the hepatic artery. These small vessels extend out from portal areas, along the boundaries between adjacent lobules. An acinus is typically diamond-shaped in cross section, with a hepatic arteriole crossing the center and with central veins at the two opposite corners. The acinus includes triangular portions of two adjacent lobules.
- There is no fundamental difference between these two concepts; they simply focus attention on different aspects of liver vasculature.
|
The Liver Lobule, from Blue Histology (Copyright Lutz Slomianka 1998-2004) (The image should be animated, if you watch patiently.)
There are other ways of dividing the parenchyma of the liver into units. Two common ways are divisions into portal lobules or into liver acini. Portal lobules emphasize the afferent blood supply and bile drainage by the vessels of the portal triads. The secretory function of the liver is emphasized by the term liver acinus. Acini are smaller units than portal or "classical" liver lobules and relate structural units to terminal branches formed by the vessels of the portal triad but not the portal triad itself. [In the animation of the acinus, the green arrows represent bile drainage.] Representations of portal lobules and liver acini vary in different textbooks. |
Hepatic Cords
The bulk of the liver consists of epithelial hepatocytes arranged into cords, separated by vascular sinusoids.
The cords of hepatocytes represent the parenchyma of the liver. Neoplasms display abnormal architecture of hepatic parenchyma.
In section, hepatic cords appear linear (hence the name, "cord"). However, these are not really strings of cells; they are more like intricately branching and interconnecting sheets, one cell thick, which extend parallel to the long axis of the lobule and radiate out from the its center.
The structural concept of epithelial cords should be contrasted with that of tubule and of acinus.
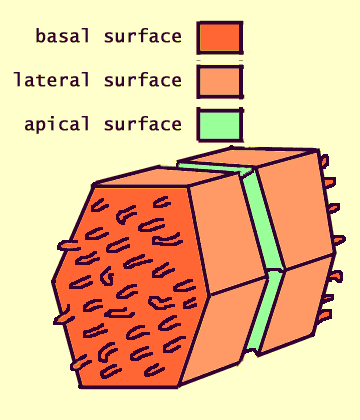
A cord consists of hepatocytes. Each hepatocyte is attached to its neighbors all around and faces the sinusoids at either end. (See hepatocytes for more detail and explanation.)
The sinusoids are vascular spaces lined by a fenestrated endothelium (i.e., an endothelium that is full of holes -- from fenestra, window). This endothelium has no underlying basement membrane. Therefore, the fenestrations permit blood plasma to wash freely over the exposed surfaces of the hepatocytes in the space of Disse.
The space between the endothelium and the cords is named the space of Disse (after Joseph Disse, b. 1852). Its location is that of connective tissue, and it does contain a network of reticular fibers (collagen type III) which hold the hepatocytes together.
More significantly, since the fenestrations of the endothelium permit free movement of blood plasma, the "interstitial fluid" of the space of Disse is blood plasma. Hence, for all practical purposes, hepatocytes reside in direct contact with blood.
Bile canaliculi, formed by apical surfaces of adjacent hepatocytes, form a network of tiny passages contained within each cord. (See hepatocytes for more detail and explanation.)
Cautionary note: Some otherwise excellent textbooks (as well as a distressingly high proportion of websites) misrepresent the three-dimensional configuration of bile canaliculi within hepatic cords. The usual error is to diagram canaliculi as if they were sandwiched between two layers of hepatocytes; in fact, each cord is only one cell thick, with a canaliculus encircling each hepatocyte.The microscopic appearance of cords and sinusoids can vary markedly depending on quality of histologic preparation.
In some specimens, sinusoidal spaces are quite evident (as in the image at left). In other specimens, sinusoidal spaces are obliterated by swollen hepatocytes (as at right).
Hepatic Sinusoids and Space of Disse
Blood from both portal vein and hepatic artery (the liver's dual blood supply) mixes together in the hepatic sinusoids and then drains out of the lobule through the central vein, a branch of the hepatic vein.
The large sinusoidal volume permits sinusoidal blood to "percolate" comparatively slowly through the liver parenchyma, thereby associating intimately with hepatocytes. This, in turn, provides time for efficient transfer of substances across the hepatocyte membrane.
Associated with the sinusoids are stellate Kupffer cells (the name commemorates Karl Wilhelm von Kupffer, b. 1829) -- liver macrophages which effectively catch and destroy bacteria which entered the blood in the intestine.
The endothelium which lines liver sinusoids is fenestrated (i.e., full of holes -- from fenestra, window) and lacks a basement membrane. The fenestrations permit blood plasma to wash freely over the exposed surfaces of the hepatocytes through the space of Disse.
The space between the fenestrated endothelium and the cords is named the space of Disse.
Historical note: In 1890, long before electron microscopy had resolved sinusoidal endothelium with its fenestrations, Joseph Disse had experimentally demonstrated the presence of the eponymous underlying plasma-filled space.
Stellate Ito cells (named after Toshio Ito), located at intervals within the space of Disse, store fat and vitamin A.
The location for the space of Disse is that of connective tissue, and, as one might thus expect, this space does contain scattered reticular fibers (collagen) and fibroblasts. More significantly, since the fenestrations of the endothelium permit free movement of blood plasma, the "interstitial fluid" of the space of Disse is blood plasma. Hence, for all practical purposes, hepatocytes reside in direct contact with blood.
Liver Pathology
(Please review the general note about pathologic histology in year one. )Because the body depends on so many various liver functions, the liver is also involved in a variety of pathological processes. As a result, many diseases give rise to characteristic changes in the appearance of the liver. The microscopic appearance of the liver can therefore be extremely useful for diagnosis.
Even advancing age can be estimated by the accumulation of lipofuscin pigment in hepatocytes.
Fortunately, the liver is easily accessible for needle biopsy (directly through the abdominal wall). It also heals readily. So microscopic examination of liver biopsy is a fairly routine clinical procedure.
Failure of adequate venous drainage from the liver leads to chronic passive congestion, with blood backing up in centrilobular areas. This gives rise to a gross appearance picturesquely labelled "nutmeg liver". For images of congested liver, see WebPath (gross) and WebPath (microscopic), or Milikowski & Berman's Color Atlas of Basic Histopathology, pp. 302.
A high concentration of leukocytes in portal connective tissue is a sign of inflammation, or hepatitis. For examples, see WebPath, or Milikowski & Berman's Color Atlas of Basic Histopathology, pp. 278-283).
Because blood flows from the periphery toward the center of each liver lobule, hepatocytes in the periphery respond first to whatever demands are placed upon them by incoming blood. Peripheral cells are therefore most vulnerable and first to be damaged by incoming insults, such as alcohol or other toxins. With sufficient liver damage, many portal areas may collapse together (portal collapse, WebPath).
Although hepatocytes are capable of considerable regeneration, chronic damage can lead to scarring as the missing hepatocytes are replaced by fibrous connective tissue. If the damaged hepatocytes are located peripherally in lobules (a common pattern, as noted above), the resulting scar expands out from adjacent portal areas.
In human liver, the appearance of connective tissue extending out from portal areas is pathological, the defining characteristic of cirrhosis. For images of cirrhosis, see WebPath or Milikowski & Berman's Color Atlas of Basic Histopathology, pp. 284-293.
Cirrhosis is a non-specific descriptive term, equivalent to "sclerosis," "fibrosis," or simply "scarring." It is indicative of a past history of tissue destruction. The associated presence of inflammatory cells suggests an ongoing process of damage.
Neoplasms display abnormal architecture of hepatic parenchyma. For examples, see WebPath (hepatocellular carcinoma), WebPath (cholangiocarcinoma), and WebPath (metastatic ductal carcinoma from breast), or Milikowski & Berman's Color Atlas of Basic Histopathology, pp. 298-301.
Clinical note: Histotripsy, the non-invasive, mechanical destruction of tissue with focused ultrasound, has recently received FDA approval for treatment of liver tumors.
|
|
|
 |
|
 |
|
|
|
|
|
 |
|
|
 |
 |
|
SIUC / School
of Medicine / Anatomy / David
King
https://histology.siu.edu/erg/liver.htm
Last updated: 21 December 2023 / dgk
2
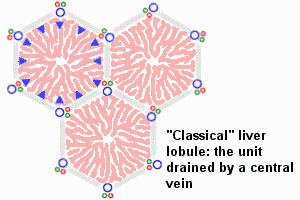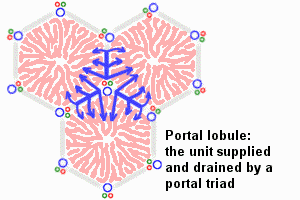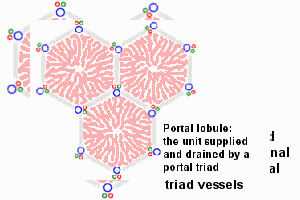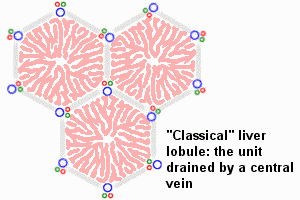 An
idealized representation of the "classical" liver lobule is a six-sided
prism about 2 mm long and 1 mm in diameter. It is delimited by interlobular
connective tissue (only little, if any, visible in humans; plentiful in
e.g. pigs). In its corners we find the portal triads. In cross
sections, the lobule is filled by cords of hepatic parenchymal cells,
hepatocytes, which radiate from the central vein and are separated by
vascular sinusoids. [In the animation of the classical and portal
lobules, the blue arrows
represent mixed portal and arterial blood.]
An
idealized representation of the "classical" liver lobule is a six-sided
prism about 2 mm long and 1 mm in diameter. It is delimited by interlobular
connective tissue (only little, if any, visible in humans; plentiful in
e.g. pigs). In its corners we find the portal triads. In cross
sections, the lobule is filled by cords of hepatic parenchymal cells,
hepatocytes, which radiate from the central vein and are separated by
vascular sinusoids. [In the animation of the classical and portal
lobules, the blue arrows
represent mixed portal and arterial blood.]