Histology Study Guide
Respiratory Tract
These notes are an ancillary resource, NOT a substitute for scheduled resource sessions or for textbooks. If you use this on-line study aid, please refer to your textbooks and atlases for richer, more detailed information.
Introduction / overview
- Naso-pharynx
Conducting system (air passageways)
Respiratory region
- Vasculature
- Pulmonary pathology (miscellaneous examples which might be instructive)
- RESPIRATORY IMAGE INDEX
Preparation
Before studying the histology of any particular system or organ, one should appreciate the basic concepts and tools of histology, as presented in the Introduction to Histology at this website. In particular, one should be familiar with the four basic tissue types, most especially epithelium and connective tissue and with the basic tools of histology.
Overview of Respiratory Tract Histology
The lung is one of several organs that packs a large epithelial surface area into a compact volume. The basic organizational pattern is that of a gland, in which a branching tree of tubes provides continuity from the body's outside surface to a vast number of epithelial cells.
- Indeed, the respiratory tract begins life as an invagination of epithelial (endodermal) tissue. Embryonic lungs even have the histological appearance of glands (see Webpath). Only fairly late in development do the cuboidal epithelial cells of the terminal alveoli assume the thin squamous epithelial shape that characterizes the lining of mature gas-exchanging air sacs. (Some significant secretory function is retained, in the form of cuboidal, surfactant-producing great alveolar cells.)
- In the respiratory system, just as in large glands, a system of conducting passageways form a branching "tree," with functional units at the end of each twig.
- In the respiratory system, the tree's "trunk" is the trachea, larger branches are called bronchi (singular "bronchus"), and smaller branches are called bronchioles. (In a gland, the conducting passages are called "ducts.")
- In the lung, the epithelial cells at the ends of all the twigs form "respiratory units," also called alveoli (singular, "alveolus"). In a gland, the secretory units at the ends of the twigs are also sometimes called "alveoli," which means a small hollow or cavity.
The pleural cavity is lined by mesothelium. This includes both the outer surface* of lung and the adjacent inner surface* of the chest wall. (Simple squamous mesothelial tissue also lines the other major body cavities, pericardial and peritoneal.)
* Note that "outer surface" in the above paragraph refers to the surface that a surgeon encounters upon opening the ribcage, while "inner surface" refers to the gas-exchange membranes "inside" the lung. These "inner" surfaces actually face inspired air, and are therefore more properly the "outer" surfaces of alveolar epithelium.
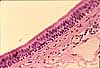
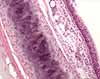
The conducting passageways of the respiratory system (nasal cavity, trachea, bronchi and bronchioles) are lined by pseudostratified columnar epithelial tissue, which is ciliated and which includes mucus-secreting goblet cells. Incoming particulates (dust, bacteria) adhere to the mucus, which is then swept upward and away by the cilia.
An extensive vascular plexus allows heat-exchange to condition air before it reaches the delicate alveoli.
Because the passage of air depends on wide open passageways, the larger respiratory passages (trachea, and bronchi) are held open by supportive skeletal elements made of cartilage.
The respiratory, gas-exchange surface consists of simple squamous epithelium lining millions of small sacs, called alveoli. Between adjacent alveoli is a network of capillaries. The epithelium lining each alveolus is exceedingly thin to facilitate diffusion of oxygen and CO2 across that lining into blood. This gas-exchange function of this pulmonary vasculature is critical to organ function and to life itself. (Alveolar epithelium also contain cuboidal surfactant-secreting cells. The surfactant overcomes the tendency of alveolar walls to adhere to one another, which would obliterate the air space).
TOP OF THIS PAGE / CONTENTS FOR THIS PAGE / RESPIRATORY IMAGE INDEX
Nasopharynx
The nasal cavity, like the rest of the larger respiratory passageways, is lined by ciliated, pseudostratified columnar epithelium with goblet cells.
The olfactory mucosa is lubricated by Bowman's glands (named after William Bowman, b. 1816).
The mucosal surfaces of the nasal passages are important for conditioning air before it reaches the more-delicate alveolar tissue. Many small glands provide moisture-containing mucus for humidification as well as for capturing incoming dust. A vascular plexus facilitates heat exchange.
The most distal expression of this heat-exchange mechanism is found in Little's area (commemorating James Little, b. 1836), located inside the nose just beneath the anteroinferior epithelial surface of the nasal septum. Also known as Kiesselbach's area (commemorating Wilhelm Kiesselbach, b. 1839), this site contains arterial anastomoses (Kiesselbach's plexus). Delicacy of the overlying epithelium makes this relatively-exposed site a frequent source of nosebleeds.Have you ever noticed what a big nose a camel has? Nasal air-conditioning is especially effective in this animal. When the creature inhales, hot dry desert air is humidified by evaporation of nasal secretions, while the nasal passages themselves are chilled by evaporative cooling. During exhalation, the cooled passages condense and recover some of the moisture, thereby contributing to water conservation.
In our own bodies, air conditioning is not as effective as that of the camel. But evaporative cooling of the respiratory passages does provide a significant contribution to thermoregulation during vigorous exercise. (In other words, many of the calories expended during exercise do not do any useful work but only generate heat, which in turn evaporates water into the air that we exhale.)
Pseudostratified respiratory epithelium begins in the nasal cavity and continues with minor variations through the nasopharynx and down the trachea, bronchi and bronchioles.
The contrast between respiratory epithelium of the nasopharynx and the stratified squamous epithelium of the oral cavity can be seen in a cross-section of the palate (or, similarly, a cross section of the epiglottis).
Research news (2026): The molecular basis for odor discrimination has long been a topic of research interest. In humans, there are hundreds of of genes encoding odor-receptor proteins, with one type of receptor per olfactory cell. The spacial distribution of differing receptor-cell types has long been presumed to be random. Researchers have recently discovered a systematic topographic arrangement of receptor types, at least in the mouse. This raises the possibility that the coding of olfactory signals to the brain shares with other sensory systems (sight, sound, touch) a topographic basis. An account of this development appeared in The New York Times on April 29, 2026. [Professional reports were published in the journal Cell by Brann et al. (2026) doi.org/10.1016/j.cell.2026.03.051 and by Bintu et al. (2026) doi.org/10.1016/j.cell.2026.03.053.]
Olfactory epithelial cells are quite wonderful, even beyond their utterly bizarre shape that displays elements of both epithelial and neural form. Renowned physician-essayist Lewis Thomas chose these cells as one of the "Seven Wonders of the Modern World":
"My Fifth Wonder is the olfactory receptor cell, located in the epithelial tissue high in the nose, sniffing the air for clues to the environment, the fragrance of friends, the smell of leaf smoke, breakfast, nighttime and bedtime, and a rose, even, it is said, the odor of sanctity. The cell that does all these things, firing off urgent messages into the deepest parts of the brain, switching on one strange unaccountable memory after another, is itself a proper brain cell, a certified neuron belonging to the brain but miles away out in the open air, nosing around the world. How it manages to make sense of what it senses, discriminating between jasmine and anything else non-jasmine with infallibility, is one of the deep secrets of neurobiology. This would be wonder enough, but there is more. This population of brain cells, unlike any other neurons of the vertebrate central nervous system, turns itself over every few weeks; cells wear out, die, and are replaced by brand-new cells rewired to the same deep centers miles back in the brain, sensing and remembering the same wonderful smells. If and when we reach an understanding of these cells and their functions, including the moods and whims under their governance, we will know a lot more about the mind than we do now, a world away." [Lewis Thomas, Seven Wonders, pp. 55-63, in Late Night Thoughts on Listening to Mahler's Ninth SymphonyViking Press, 1983.]
Consult your histology textbook and/or atlas (e.g., Rhodin, figures 31-5 to 31-8) for additional detail and electron micrographs of olfactory cells.
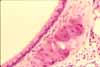
The pharynx houses tonsils, which are localized lymphoid specializations located in the mucosa not only of the pharynx but also of the tongue and the palate. Each tonsil consists of an epithelial crypt (an invaginated pocket) surrounded by dense clusters of lymph nodules.
Lymph nodules (also called lymphoid follicles) are sites where lymphocytes congregate. At the center of each lymph nodule is a "germinal center" where the lymphocytes proliferate.
Pharyngeal tonsils (also called "adenoids") provide immune surveillance cells (lymphocytes) with sites for encountering foreign antigens which enter the body through inspired air.
For more on Mucosa-Associated Lymphoid Tissues (MALT), consult your histology and immunology texts.
Pharyngeal tonsils resemble those of the palate (palatine tonsils, thumbnail to right), but with ciliated, pseudostratified columnar epithelium rather than stratified squamous epithelium lining the surface and crypts.
TOP OF THIS PAGE / CONTENTS FOR THIS PAGE / RESPIRATORY IMAGE INDEX
Conducting system.
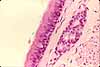
Most of the larger respiratory passageways, from the nasal cavity through the bronchi, are lined by a pseudostratified columnar epithelium.
This respiratory epithelium consists primarily of columnar ciliated cells interspersed with mucus-secreting goblet cells. Ciliary beating sweeps mucus and the dust it carries up the bronchi and trachea toward the pharynx, where it can be swallowed.
Further down the tree, the columnar epithelium of trachea and bronchi transitions to simple cuboidal epithelium of bronchioles. The height of this epithelium decreases as one descends bronchiolar branches toward gas-exchange regions. (Lung alveoli, in contrast, are lined by very thin simple squamous epithelium.)
Electron micrographs of cilia may be seen at WebPath and at the Electron Microscopic Atlas.
(A ciliated epithelium is also characteristic the female reproductive tract.)
Scattered among the ciliated cells are occasional mucus-secreting goblet cells. Mucus moistens the epithelial surface (so the ciliated cells can function) and adheres to inhaled particles (so dust and bacteria are caught before they are carried all the way into alveoli).
For additional detail on the properties of respiratory mucous in relation to ciliary function, see B. F. Dickey (2012) Walking on Solid Ground, Science, 24 August 2012: 924-925..
The name "goblet cell" refers to the cell's shape: narrow at the base and bulging apically. The apical end of each goblet cell is occupied by a large mass of mucus, which compresses adjacent cells (thus conferring the characteristic "goblet" shape) and displaces the nucleus toward the basal end of the cell. As in other mucous cells, the nucleus is compact and intensely stained (basophilic) with H&E. (Mucus-secreting cells of similar shape, also called "goblet cells," may be found in the gastrointestinal tract.)
Basal cells (those whose nuclei lie low in the epithelium, near the basement membrane) are believed to be the source of replacement ciliated and goblet cells.
Consult your histology textbook and/or atlas (e.g., Rhodin, figure 31-22) for additional detail and electron micrographs of these cells.
TOP OF THIS PAGE / CONTENTS FOR THIS PAGE / RESPIRATORY IMAGE INDEX
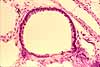
Trachea, bronchi, and bronchioles
The trachea is the "trunk" of the branching tree of passageways leading into the lungs. Its major histological specializations include skeletal reinforcement in the form of incomplete (i.e., C-shaped) rings of cartilage and a vascular (venous) plexus which facilitates heat exchange to condition air before it reaches the delicate alveoli.
For details of the ciliated, pseudostratified epithelium of the trachea, see above.
The trachea divides into two primary bronchi (singular, bronchus), one to each lung. The primary bronchi then divide repeatedly into smaller bronchi and even smaller bronchioles. Thus bronchi and bronchioles are, respectively, the larger and smaller branches of the air-conducting tree extending into the lung.
Several parameters -- the amount of cartilage, the thickness of the respiratory epithelium, and the degree of pseudostratification -- all decrease as one descends the air-conducting passages toward the alveoli.
The distinction between bronchi and bronchioles is somewhat arbitrary, but in general bronchi have cartilagenous reinforcement and a columnar epithelial lining, while bronchioles lack cartilage and have a simple cuboidal lining.
Besides ciliated and mucus-secreting cells (see above), bronchiolar epithelium may include club cells (bronchiolar exocrine cells) whose role remains incompletely understood but which seem to have some specialized synthetic and secretory function.
Research review: Rokicki, Wojciech et al. (2016), "The role and importance of club cells (Clara cells) in the pathogenesis of some respiratory diseases," Kardiochirurgia i torakochirurgia polska = Polish journal of cardio-thoracic surgery, vol. 13, pp. 26-30. doi:10.5114/kitp.2016.58961.
Historical note: Older textbooks refer to bronchiolar exocrine cells as "Clara cells," a name which recognizes researcher Max Clara (b. 1899) who worked on bodies of prisoners executed by the Nazi regime prior to WWII (e.g. see here). Because of this tainted history, the alternative term "club cells" has been adopted by several journals and societies.
Small mucous glands are scattered along most of the respiratory tree and may occasionally be seen in the wall of bronchi or bronchioles.
The airway walls also feature smooth muscle, which permits regulation of the distribution of air through the lung's volume.
TOP OF THIS PAGE / CONTENTS FOR THIS PAGE / RESPIRATORY IMAGE INDEX
Respiratory (gas-exchange) region of lung
The lung consists of functional respiratory units called "air sacs" or alveoli (singular, alveolus),
Because the actual shape of the gas-exchange spaces is somewhat complex, additional descriptive terminology is sometimes used. Terminal bronchioles, those which open directly into gas-exchange spaces, are called respiratory bronchioles. These typically open into passageways, called respiratory ducts, that open not only into alveoli but also into further passageways, called respiratory sacs, that themselves open into more alveoli.
In elementary science classes, the lung is sometimes misleadingly portrayed as a balloon at the end of a large bronchus. There is no balloon. One might imagine that each alveolus is a very tiny balloon, but even this is not accurate. The lung is more like a well-organized sponge. Each alveolus shares its wall (inter-alveolar septum) with adjacent alveoli. Alveoli cannot collapse individually, only all together.Note on clinical histology: The lung's normal volume is mostly air-filled space. In ideal histological specimens, this space is preserved by keeping the lung inflated during preparation. But as a matter of convenience, clinical specimens are often prepared in a somewhat collapsed state. Do not be surprised by lung specimens in which alveoli appear flattened, with alveolar walls wrinkled and pressed together, with reduction in the air-space.

Alveoli are separated from one another by thin alveolar walls (interalveolar septa). Each alveolus is lined by simple squamous epithelium, exceedingly thin to facilitate diffusion of oxygen while still forming an epithelial barrier between the outside air and the internal body fluids.
In between the epithelial surfaces of adjacent alveoli lies a network of capillaries, through which flow the red blood cells whose hemoglobin absorbs oxygen for transport to the rest of the body (also see lung vasculature, below).
In other words, each interalveolar septum consists of simple squamous epithelium on either side with capillaries sandwiched in between.
Before being absorbed by hemoglobin, an oxygen molecule must first reach an alveolus, then diffuse across the plasma membrane of a squamous epithelial cell, through the very thin epithelial cell cytoplasm, across the plasma membrane on the other side of the epithelial cell, across the basement membrane which separates the epithelium from the underlying tissue, across the basement membrane which surrounds each capillary (these two basement membranes may be fused together), across the plasma membrane of a capillary endothelial cell, through the very thin cytoplasm of this cell, across the plasma membrane on the capillary-lumen side of the endothelial cell, through a bit of blood plasma, and finally across the membrane of a red blood cell. Whew! The total thickness may be less than 0.05 μm.
An electron micrograph of the blood-air barrier may be seen at the Electron Microscopic Atlas.
Clinical note: Alveoli can provide a cozy haven for bacterial growth, leading to pneumonia and inflammation. For an image of alveoli filled with inflammatory cells, see WebPath. (For additional lung pathology, see below.)
The squamous epithelial cells of the alveolar walls are sometimes called Type I pneumocytes. As noted above, these squamous Type I cells facilitate gas exchange.
Scattered among the squamous cells are occasional larger cuboidal cells, sometimes called greater alveolar cells and sometimes called Type II pneumocytes. These cells secrete surfactant, which serves the critical function of keeping alveolar walls from sticking to one another.
Consult your histology textbook and/or atlas (e.g., Rhodin, figures 31-45 to 31-54) for additional detail and electron micrographs of cells associated with lung alveoli.
See the Electron Microscopic Atlas for (mostly unlabelled) EM images of lung.
The edges of inter-alveolar septa, at the entry-ways into respiratory sacs and alveoli, contain small bundles of smooth muscle which allow some control of air distribution among various regions of the lung.
To keep the alveoli clean, amoeboid phagocytic cells crawl over the surface of the alveolar walls and ingest any foreign material that they encounter. These cells are sometimes called "dust cells," but they are basically alveolar macrophages that have moved out of connective tissue and onto the epithelial surface of the alveoli.
It is perhaps amusing to note that these cells are behaving rather like independent agents, having left the body proper by crossing the alveolar epithelium. Although they may again cross the epithelium and re-enter the body, these cells may also be swept up the airway by cilia and swallowed, in effect becoming food.
TOP OF THIS PAGE / CONTENTS FOR THIS PAGE / RESPIRATORY IMAGE INDEX
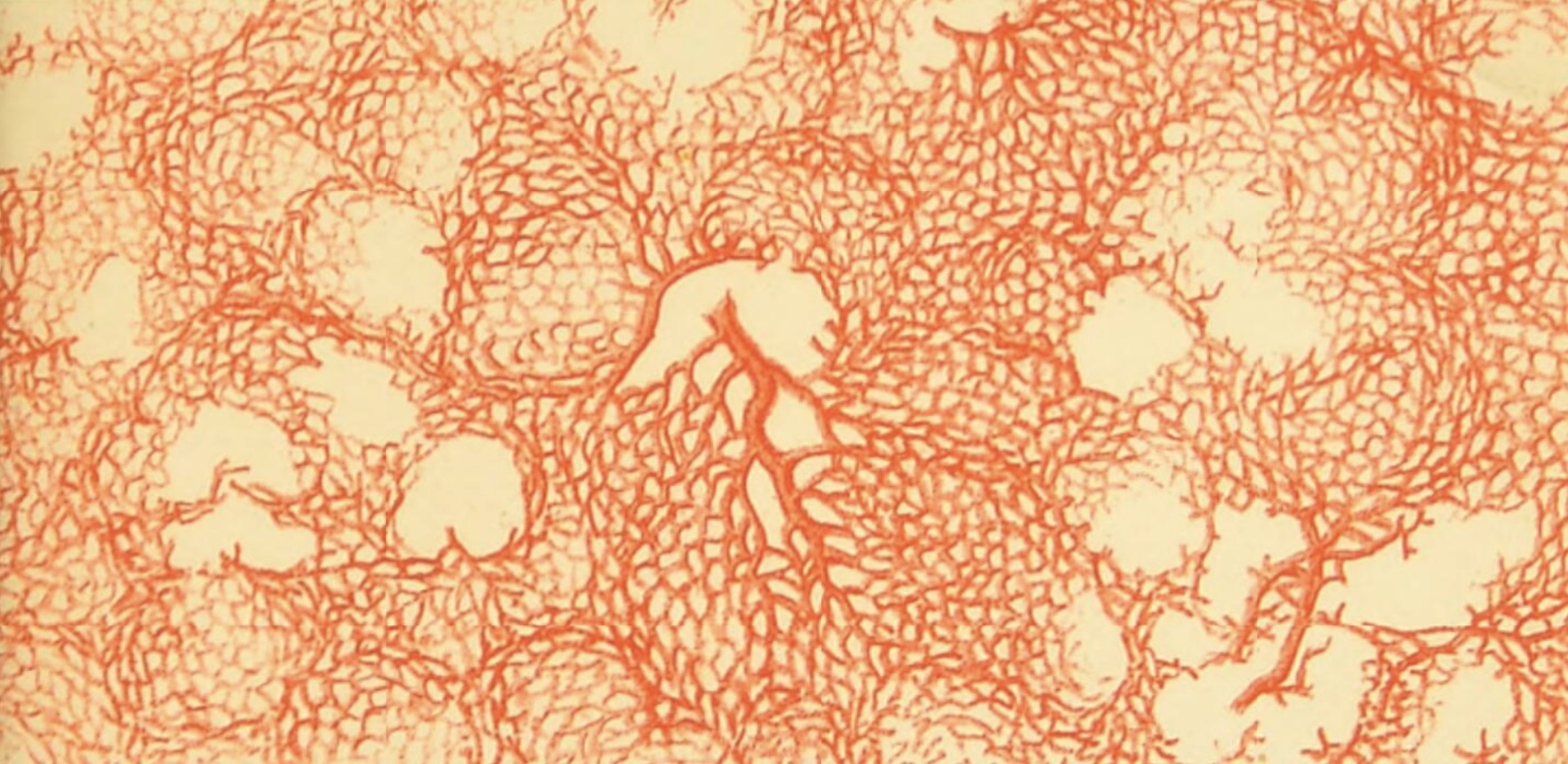
Lung vasculature
Because pulmonary circulation has lower pressure than systemic circulation, pulmonary arteries and veins tend to have more delicate walls than those seen elsewhere in the body.
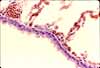
Historial note: William Harvey (b. 1578), who first understood the circulation of blood, lacked the means to directly observe the capillary connections between artieries and veins. Capillaries were first reported a few years after Harvey's death, by Marcello Malpighi (b. 1628), based on his microscopic observations of frog lung.
Consult your histology textbook and/or atlas (e.g., Rhodin, figures 31-55 to 31-67) for additional detail and electron micrographs of pulmonary vessels.
TOP OF THIS PAGE / CONTENTS FOR THIS PAGE / RESPIRATORY IMAGE INDEX
Examples of pulmonary pathology,
The following links illustrate some departures from normal lung-tissue appearance, presented at the WebPath website. In these images, try to recognize the basic features of lung tissue as well as the particular pathology.
- Pneumonia. Alveoli become filled with inflammatory cells
- Pneumonia, another example.
- Absessing pneumonia. Inflammation with tissue destruction.
- Bronchial asthma. Bronchial walls undergo various changes.
- Fat embolism. Adipocytes clog respiratory blood arterioles following trauma that damages marrow of bones.
- Tuberculosis. Granulomas form in lung tissue.
- Cancer. A malignant neoplasm metastasizes to lung.
- Disseminated intravascular coagulation.
- Pulmonary thromboembolus (animated diagram).
- Pulmonary embolus.
TOP OF THIS PAGE / CONTENTS FOR THIS PAGE
Image index
 Comments
and questions: dgking@siu.edu
Comments
and questions: dgking@siu.edu
SIUC / School
of Medicine / Anatomy / David
King
https://histology.siu.edu/crr/rsguide.htm
Last updated: 1 May 2026 / dgk