Introduction to Skin Histology
The skin is the largest organ of the body. As the primary interface between ourselves and our environment, the skin serves several distinct functions.
- protection
- sensation
- thermoregulation
- communication.
- Skin is also self-repairing after injury.
The microscopic anatomy of skin reflects this functional complexity, with each functional specialization implemented by particular features of cell and tissue structure.
- LAYERS of skin
- APPEARANCE of skin
- CELL TYPES
- Epidermal cells -- keratinocytes, melanocytes, Langerhans cells, Merkel cells
- Dermal cells -- see connective tissue cells
- SPECIAL STRUCTURES
- REGIONAL DIFFERENTIATION
- FUNCTIONS of skin

- Containment
- Protection
- Healing after injury
- Sensation
- Thermoregulation
- Communication
- BIOPSY -- Examine and interpret a skin biopsy.
- Recent research: A special issue of the journal Science (21 November 2014, Vol. 346 no. 6212) reports research progress on several skin-related topics.
- "From bench to bedside," Science 346: 932-933
- "Shedding light on skin color," Science 346: 934-936
- "Mammalian skin cell biology: At the interface between laboratory and clinic," Science 346: 937-940
- "Advances in skin grafting and treatment of cutaneous wounds," Science 346: 941-945
- "The melanoma revolution: From UV carcinogenesis to a new era in therapeutics," Science 346: 945-949
- "The gentle touch receptors of mammalian skin," Science 346: 950-54
- "Dialogue between skin microbiota and immunity," Science 346: 954-959
- SAQ self-assessment practice questions
- Image index
- Link to LUMEN (Loyola University Medical Education Network), click on "Part 13: Skin (Integument)".
The skin has two principal layers.
- The epidermis is the epithelial tissue layer of skin.
- Hair follicles, sebaceous glands, and sweat glands are epithelial invaginations from the epidermis.
dermis is the connective tissue layer of skin.
- Embedded within the dermis are blood vessels and sensory nerve endings as well as epidermal invaginations of hair follicles and sweat glands.
Beneath the skin lies the hypodermis -- connective tissue which may be adipose or fibrous, depending on location.
- The transition from dermis to hypodermis is irregular and poorly defined; there is no "boundary".
The quality of the epidermis differs from place to place in the body (see regional differences). The quality of the epidermis can also be altered by various disease states which influence the rate of cell division and the quality of cell differentiation.
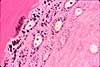
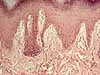
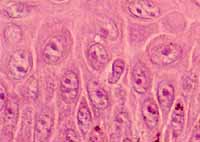
The epidermis displays several layers. These layers are not distinctly different tissues (unlike epidermis and dermis, for example) but rather reflect visible changes or stages along the continuous process of keratinocyte maturation, or keratinization. Read the caption of the image below from the bottom up, for the normal progression of keratinocytes from formation to maturation.
 |
- | |
 |
- |
Stratum granulosum.
Cells in the granule-cell layer accumulate keratohyalin,
visible as darkly stained granules. The presence of this layer is
diagnostic for keratinized stratified
squamous epithelium.
|
 |
- |
Stratum spinosum.
Cells of the "prickle-cell" layer are attached to one
another by desmosomes ("spines") and reinforced by tonofilaments.
These cells gradually move outward as new cells are formed from
the basal layer (below).
|
 |
- | |
 |
- |
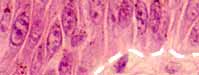
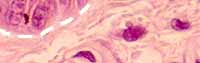
The dermis
lies beneath the epidermis, separated from the epithelium by the basement
membrane (white dashed line).
|
Keratinocytes
The epidermis consists primarily of keratinocytes. Scattered among the keratinocytes are a few other cell types -- melanocytes, Langerhans cells, and Merkel cells
Keratinocytes, which comprise most of the epidermis, are characterized by numerous intercellular junctions (desmosomes), reinforced by intracytoplasmic tonofilaments.
Each desmosome is one spot of attachment. At high magnification, the desmosomes are visible as fine "prickles" extending across the gap (intercellular space) between adjacent keratinocytes. Between these junctions lie intercellular channels which permit nutrients to diffuse from dermis into epidermis. (More.)
Keratinocytes in the stratum basale of the epidermis can undergo mitosis. The formation of new cells in this basal layer gradually pushes previously formed cells upward where they become the stratum spinosum. As keratinocytes approach the surface of the epidermis, they accumulate intracellular keratin and secrete a waxy material into the intercellular space; these changes are visible in the stratum granulosum, a distinctive layer which is diagnostic for a keratinized epithelium. As maturing keratinocytes seal off the intercellular spaces through which they receive nutrients, they eventually die and form the stratum corneum, a tough and relatively inpermeable layer of hardened, dead cells. Eventually, as cells reach the surface, they are sloughed off. The entire epidermis above the basal layer is replenished (replaced by new cells) within about two weeks. Replacement is accelerated by injury.
The stages in keratinocyte maturation appear as layers in the epidermis, so that a section across the epidermis illustrates the entire process.
See the Electron Microscopic Atlas of cells, tissues and organs in the internet for (mostly unlabelled) EM images of epidermis.
Other epidermal cell types
Scattered among the much more numerous keratinocytes are several other epidermal cell types -- melanocytes, Langerhans cells, and Merkel cells. Because these cells lack the tough reinforcement and desmosomal attachments that characterize keratinocytes, they commonly shrink during preparation and appear surrounded by a clear "halo". (Together these cell types are all quite distinct from keratincytes. But they are difficult to distinguish from one another without special techniques.)
Melanocytes manufacture the pigment melanin. Melanocytes appear as small cells, usually in or near the stratum basale. They have thin cytoplasmic processes (not evident in ordinary histological preparations) which extend between nearby keratinocytes and serve to transfer melanosomes (melanin-containing granules) into adjacent keratinocytes. Because of this transfer, most pigment-containing cells in the epidermis are normally keratinocytes rather than melanocytes.
Melanocytes may be found in places other than skin, such the choroid layer of the eye.
Melanocytes are derived from neural crest and migrate to their final position in the epidermis. This developmental propensity for travel may contribute to the dangerously metastatic potential of melanomas.
- Recent research: "The melanoma revolution: From UV carcinogenesis to a new era in therapeutics," Science 346: 945-949 (2014)
Langerhans cells (named after Paul Langerhans, b. 1847) are antigen-presenting cells which participate in the surveillance function of the immune system. (Antigen-presenting cells acquire foreign materials [antigens] and pass them along to lymphocytes.) Langerhans cells are smaller than keratinocytes, with relatively clear cytoplasm, usually located within the stratum spinosum or stratum basale. Langerhans cells are dendritic cells, with extensive cytoplasmic processes extending between keratinocytes to sample intruding antigens throughout the epidermis. [NOTE: Please try not to confuse Langerhans with Langhans, each of whom has an eponymous cell named after him.]
Merkel cells (named after Friedrich Merkel, b. 1845) are small cells associated with nerve endings in epidermis. Their function has long been uncertain, but they seem to be involved in neural development and tactile sensation. Recent evidence supports a role for Merkel cells in light touch, "suggesting that these cells form an indispensible part of the somatosensory apparatus" (Science 324:1580, 2009; also see The Journal of Neuroscience 32(10): 3296-3300, doi: 10.1523/JNEUROSCI.5307-11.2012 ). Some uncommon skin cancers derive from Merkel cells.
See the Electron Microscopic Atlas of cells, tissues and organs in the internet for (mostly unlabelled) EM images of Merkel cells.
Dermis and hypodermis
The dermis consists of dense, fibrous connective tissue whose predominant connective tissue component is collagen.
- The texture of collagen fibers serves as the basis for recognizing two layers of dermis.
- The papillary layer of the dermis lies adjacent to the epidermis and consists of relatively small, finely textured collagen fibers. This layer is named after dermal papillae, the protrusions of dermal connective tissue which indent the base of the epidermis. Dermal papillae increase the contact area for attachment between dermis and epidermis; they are more pronounced in thick skin.
- The reticular layer of the dermis lies beneath the papillary layer and consists of larger, more coarsely textured collagen fibers. ("Reticular" means "like a network" and describes the texture of collagen fibers in this layer.)
Like ordinary connective tissue throughout the body, connective tissue of the dermis serves several distinct functions.
- Tough collagen fibers and resilient elastic fibers provide mechanical strength for skin.
Clinical note: Lines of tension in the dermis, called Langer's lines (after Karl Langer, b. 1817), affect healing after surgical incision. Cuts across the lines tend to pull apart, with concommitant tendency toward tissue distortion and scarring, more so than do cuts parallel to the lines.
- The ground substance of the dermis serves as the substrate for diffusion of nutrients and wastes to and from various other tissue components.
Mast cells, lymphocytes and macrophages in the connective tissue carry out surveillance for the immune system.
- Finally, the dermis together with its associated blood vessels and nerves is capable of active response to injury, yielding the defensive reaction of inflammation, followed by the healing processes of growth and repair.
Within the dermis are embedded several other structures, including epidermal appendages (sweat glands and hair follicles) as well as blood vessels and nerve endings.
The connective tissue of the dermis grades into hypodermis, without a sharp transition or distinct boundary.
Over most of the body, hypodermis is characterized by adipocytes and may comprise a thick layer of adipose tissue. In some sites (e.g., "dimples"), hypodermis is fibrous and binds the dermis to underlying structures. Hair follicles and sweat glands may extend into hypodermis
Blood vessels are generally larger in the deeper layers of skin, with only capillaries in the papillary layer of the dermis.
The appearance of the skin can have considerable clinical significance. The skin is readily accessible for examination (no invasive procedures needed), and its color and texture can reveal much about underlying physiology.
Color: Skin is moderately transparent. Light which penetrates the skin is reflected back from varying depths by epidermal cells, by collagen, and by blood.
Recent research: "Shedding light on skin color," Science 346: 934-936
Melanin, produced by melanocytes and stored in basal keratinocytes, contributes a yellow/brown color to the epidermis. If the epidermis is not heavily pigmented, light readily penetrates into the dermis.
Collagen scatters light from the dermis without altering its color. Hence, the whiteness of "white" skin is primarily a reflection of collagen.
Hemoglobin in red blood cells scatters red light and is responsible for the pinkness of unpigmented skin. The relative amount of pink in any given patch of skin reflects how closely blood approaches the base of the epidermis (i.e., how much collagen intervenes to scatter white light before red blood cells can absorb the non-red colors).
Each of these elements contributes to the apparent color of skin. Variations in skin color in different parts of the body (see regional differences) are based on variations in these elements, most especially the amount of pigment, the thickness of dermis, and the degree of perfusion in dermal capillaries.
Perhaps most significantly, blood flow through the dermis is highly variable and is regulated in response to many conditions (heat, pain, fluid balance, inflammation, emotional reaction). Resulting variations in pinkness can provide indicators of underlying physiology, both locally and systemically. Obvious examples include inflammation, overheating, dehydration, shock, and even embarrassment (i.e., blushing) .
Texture: Skin texture is affected the thickness and smoothness of the epidermis, by the quality of fibers in the dermis, and by the amount of fluid in dermal connective tissue.
Because the epidermis is continually being replenished by cell divisions among basal keratinocytes and because this tissue is exposed to a variety of insults, the epidermis is especially prone to disturbances of growth. See any pathology book for examples.
The connective tissue fibers of the skin are permanent, enduring without replacement (except by repair after injury) throughout life. Although collagen is quite durable, elastin commonly deteriorates with age (and especially with repeated exposure to sunlight) and loses its elasticity. This is easily demonstrated by a "pinch test." In youthful skin, loose skin that has been pinched into a ridge quickly returns to its normal position when released. Elderly skin commonly remains in its deformed position, returning more slowly if at all.
Both edema (accumulation of excess fluid in connective tissue) and dehydration can dramatically alter the appearance of skin.
Skin includes several specialized structures, including epidermal appendages (sweat glands, hair follicles, nails) as well as blood vessels and nerve endings which travel through the dermis.
Epidermal appendages play an especially important role in recovery from superficial scrapes and burns. Even when the epidermis has been removed over a fairly large area, it can grow back quickly from the epithelial cells which remain in deeper hair follicles and/or sweat glands. Third-degree burns are so serious precisely because tissue damage extends deep enough into the dermis to destroy these sources of replacement cells.
Sweat glands
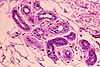
Sweat glands are simple tubular glands lined by cuboidal epithelium. The secretory portion of the gland lies deep in the dermis, where the tubule is twisted into a fairly compact tangle. A duct communicates outward through the overlying dermis and the epidermis.
The secretory portion of a sweat gland is comprised of cells which are larger than those of the duct. These cells form a simple cuboidal epithelium, along with interspersed myoepithelial cells (which can expel sweat by contraction).
Cells comprising the duct, or conducting portion of the tubule, usually form a two-layered stratified cuboidal epithelium. These cells are usually stained more intensely than those comprising the secretory portion of the tubule. As fluid flows through the duct, its composition is modified by reabsorption of certain elements from the fluid. (This is primarily a means of conserving salt.)
Sweat glands are vital for thermoregulation. They also influence water and ion balance.
The primary function for sweating is evaporative cooling of the body. Thus, the amount of sweat is regulated as a function of body temperature.
However, sweat also contains salt. Normally, sweat which comes out on the surface of the skin has a lower salt concentration than the precursor fluid produced by the secretory cells of the sweat gland. Salt is reabsorbed by the duct of the sweat gland. The effectiveness of this salt reabsorption is regulated by aldosterone (the hormone responsible for maintaining electrolyte homeostasis) in response to bodily salt balance.
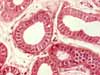
There are two types of sweat glands. Ordinary eccrine sweat glands are found over most of the body, while larger apocrine sweat glands are found in axillary, pubic, and perianal regions.
Both types of sweat glands have the same basic shape, but apocrine glands have taller cells and much larger diameter.
Hair follicles
Hair follicles are tubular invaginations lined by stratified squamous epithelium similar to epidermis.
Toward the bottom of each follicle, processes of cell division, growth, and maturation similar to those in the epidermis yield a cylindrical column of dead, keratinized cells (the hair shaft) which gradually extrudes from the follicle. (For details, consult your histology textbook.)
Hair follicles are associated with sebaceous glands as well as nerve endings and smooth muscle, which all together form the pilosebaceous apparatus.
- A network of nerve endings detects deflection of the hair shaft and also controls piloerection (hair "standing on end," or "goose bumps").
- Piloerection is effected by smooth muscle. A small bundle of smooth muscle cells called the arrector pili ("hair erector") is attached to the connective tissue sheath around each hair follicle.
- Sebaceous glands secrete oil into the hair follicle.
Sebaceous glands
Sebaceous glands are associated with hair follicles. The complex of hair follicle, hair shaft, and sebaceous gland is sometimes called the pilosebaceous apparatus.
Histologically, sebaceous glands are quite different from all other glands. They are holocrine glands, which means that the whole cell is secreted. The process of holocrine secretion is more similar to maturation of keratinocytes than to ordinary glandular function. Cells formed by mitosis at the base of the gland are pushed toward the surface as new cells form below. Along the way, the cells become packed with lipid and then die. The secretion consists of breakdown-products of the cells themselves, which extrude into the lumen of the associated hair follicle. So, basically, sebaceous glands are small masses of epidermal cells in which sebum (a mixture of lipids) accumulates rather than keratin.
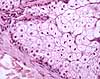
The dying cells in sebaceous glands provide a good opportunity to learn the appearance of pyknotic nuclei, one of the more conspicuous signs of cell death.
Nails
Please consult an in-depth text (e.g., Chapter 3, Histology for Pathologists, Sternberg, 1998; newer edition: Mills, Histology for Pathologists, 3rd ed., 2007) if you desire histological details on fingernails and toenails.
Innervation
The skin is richly innervated, served by a variety of sensory nerve endings which respond to a variety of modalities (e.g., pressure, vibration, heat, cold, itch, pain) and by motor nerve endings which control blood flow, sweat secretion, and piloerection.
- Recent research: "The gentle touch receptors of mammalian skin," Science 346: 950-54.
Meissner's corpuscles
Pacinian corpuscle For richer information on the following, see Neuroscience Online, Somatosensory systems.
- Free nerve endings (without any conspicuous associated structure) terminate within the epidermis, penetrating almost to the stratum corneum.
- Merkel's touch corpuscles (named after Friedrich Merkel, b. 1845) are nerve endings associated with Merkel cells at the base of the epidermis in thick (glabrous) skin of palms and soles.
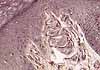
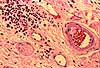
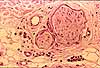
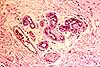
- Meissner's corpuscles (named after Georg Meissner, b. 1829) (images at right) are encapsulated endings in dermal papillae, most common in palmar and plantar skin, especially in fingertips.
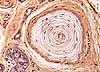
- Pacinian corpuscles (named after Filipo Pacini, b. 1812), located deeper in dermis (image at right), are simple nerve endings but are each encapsulated by multilamellar, ovoid structures resembling small onions. Pacinian corpuscles respond to deep pressure.
- Krause's endbulbs (named after Wilhelm Krause, b. 1833), or mucocutaneous receptors, are encapsulated endings in dermis, especially associated with lips, genital regions, nipples, and conjunctiva.
- Ruffini endings (named after Angelo Ruffini, b. 1864) have numerous fine branches from a single axon within the fluid-filled space of a single thin capsule.
- Hair follicle receptors are unencapsulated nerve endings wrapped around hair follicles.
The distribution of sensory nerve endings varies from place to place in the body (see regional differences).
Except for the characteristic capsules of Meissner's and Pacinian corpuscles, nerve endings are inconspicuous in ordinary histological preparations of skin.
Special stains are generally used to observe nerve endings. And except for the conspicuously encapsulated endings of Meissner's and Pacinian corpuscles, the functional details of most sensory endings remain obscure. For more information on tactile sensation, see Principles of Neural Science by Kandel, Schwartz and Jessel.
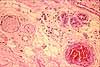
Peripheral nerves (i.e., bundles of axons, within a connective tissue sheath or epineurium) can often be found in dermis, with smaller branches toward the surface (i.e., often near sweat glands or hair follicles) and larger branches in deeper layers (often running parallel to blood vessels). The following examples show nerves in dermis.
Skin vasculature
The papillary layer of the dermis is richly supplied with capillaries, while larger blood vessels may be found in deeper levels of the dermis.
Since the skin does not have a very high metabolic demand for nutrients and oxygen, this rich vascular network serves mainly for regulation of body temperature. Essentially, regulation of the amount of blood flowing through superficial capillaries allows for either conservation or dissipation of body heat.
Arteriovenous shunts, controlled by associated sphincters, allow blood to bypass capillaries and flow directly from arteries into veins. These shunts occur in both deep and superficial dermis.
Regional DifferentiationSkin varies markedly over different parts of the body. All of the components of skin contribute to this variation. Consult a textbook for illustrations (e.g., pp. 42-43 in Histology for Pathologists, Sternberg, 1998; newer edition: Mills, Histology for Pathologists, 3rd ed., 2007).
- Thickness of epidermis.
- Skin on palms of hands and soles of feet is traditionally called "thick skin" or "volar skin." This palmar and plantar skin has much thicker epidermis than other regions of skin (up to a millimeter or more, with many cell layers), and with an especially well-developed, abrasion-resistant stratum corneum.
This so-called "thick skin" also lacks hair follicles and sebaceous glands.- Elsewhere epidermis is substantially thinner than palms and soles, typically with only a few cell layers. Nonetheless, its thickness varies from region to region -- e.g., commonly about a half-millimeter over most of the body, but as thin as a tenth of a millimeter over eyelids.
Clinical / research note (from Science, 5 Sept. 2024, Vol 385, pp. 1047-1048). "Differences between skin in different parts of the body are well recognized, but exploiting those differences to benefit the millions of people worldwide with prosthetic limbs is a new prospect. The skin of the palms and soles, known as volar skin, is specialized to withstand physical and mechanical forces, such as friction, shear stress, and pressure. Limb prostheses come into close contact with stump skin that is not adapted to these forces. As a result, the skin can break down, resulting in pain, ulceration, and infection... [Researchers demonstrated], in a clinical trial of healthy volunteers, that injecting autologous volar fibroblasts (derived from the volunteers' own tissue) confers volar features on nonvolar skin that persist for several months. This is a promising step toward improved quality of life for prosthesis wearers."- Thickness of dermis.
- Dermis is commonly one to two millimeters in thickness.
- Dermis is quite thin in the eyelid (about half a millimeter) and quite thick (several millimeters) over the back of the trunk.
- Dermal papillae are most pronounced beneath the epidermis of thick skin.
- Size and concentration of hair follicles / hair shafts.
- Rather obviously, hairs are commonly thicker and longer on the scalp than on most other regions, and in adults also on axilla and pubis.
- Less obviously, tiny hair (vellus hair) occurs even on seemingly hairless regions like eyelids.
- Hair is absent from "thick skin" of palmar and plantar skin.
- There is considerable variation from person to person in the distribution of hair.
- Size and concentration of sebaceous glands.
- Certain regions of the body (e.g., nose, forehead) are notorious for large and active sebaceous glands.
- Sebaceous glands are absent from "thick skin" of palmar and plantar skin.
- Size and concentration of sweat glands.
- The distribution of sweat glands varies over the body, with high concentrations in palmar and plantar skin. (Incidently, sweat glands of palms and soles respond more to mental and emotional stress than to heat stress.)
- Large apocrine sweat glands are concentrated in axillary, pubic and perianal areas. These sweat glands are responsible for "body odor," by including organic substances (and, consequently, bacteria) in their secretions.
- Type and concentration of sensory nerve endings.
- Different sensory modalities are concentrated in different regions.
- Skin of fingertips has the highest concentrations of Meissner's and Pacinian corpuscles.
- Tactile resolution varies tremendously from region to region, as can be readily demonstrated by a two-point discrimination test:
(Unbend a paperclip so that the two ends can be pressed simultaneously against the skin. Then, with randomly varying touches from one or both ends, see how far apart the ends need to be before the two-end touch is felt as two distinct touches. The finest discrimination is likely to be found on fingertips; the coarsest on the back of the trunk.)- Presence of muscle.
- Bundles of smooth muscle may be found in the dermis of nipple, areola, scrotum, penis, and perianal region.
- In non-human mammals, skeletal-type muscle may be found in dermis (allowing horses, for example, to "twitch" a patch of skin to discourage biting flies).
Functions of Skin
Skin serves several functions simultaneously.
- Although the stratum spinosum is permeable to water, the epidermis becomes relatively impermeable in the stratum granulosum and stratum corneum.
- Damage to extensive areas of epidermis, e.g. by burns, renders the skin highly permeable and constitutes a medical emergency.
- Epidermis serves as a simple mechanical barrier. This is probably the most obvious function for skin.
- Keratinocytes are crucial for the barrier function, both in their tonofilaments and desmosomes which establish the mechanical integrity of the epidermis and in their formation of hardened squames in the stratum corneum.
- Melanocytes produce melanin pigment, which shields underlying cells from ultraviolet light.
Recent research: "The melanoma revolution: From UV carcinogenesis to a new era in therapeutics," Science 346: 945-949; "Shedding light on skin color," Science 346: 934-936- Collagen of the dermis provides main strength to resist tearing or penetration. The thickness of the dermis is correlated with vulnerability to injury.
Immunological surveillance and defense -- Immune cells of skin stand ready to defend against invasion by microorganisms.
- Langerhans cells detect foreign antigens in the epidermis.
- Mast cells stand ready to trigger an inflammatory response if the skin is injured or the epidermal barrier is breached.
Recent research: "Dialogue between skin microbiota and immunity," Science 346: 954-959.
Cells in the basal layer of the epidermis respond quickly to damage, proliferating and migrating to cover the site of injury (moving in under the scab).
- Epithelial replacement can spread from deep hair follicles and sweat glands if the surface epidermis has been damaged over an extensive area.
- Fibroblasts also become activated by injury, to proliferate and to manufacture new collagen. The resulting scar may be eventually remodelled into a nearly-normal configuration of fibers.
- Epidermal appendages play an especially important role in recovery from superficial scrapes and burns. Even when the epidermis has been removed over a fairly large area, it can grow back quickly from the epithelial cells which remain in deeper hair follicles and/or sweat glands. Third-degree burns are so serious precisely because tissue damage extends deep enough into the dermis to destroy these sources of replacement cells.
Recent research: "Advances in skin grafting and treatment of cutaneous wounds," Science 346: 941-945.
- Sweat glands are vital for thermoregulation. Even when the external temperature is higher than body temperature, evaporating sweat can cool skin below the core temperature. Normal thermoregulatory sweating is often insensible, with sweat evaporating as quickly as it forms. Sweat which drips from the body (e.g., during heavy exercise in humid weather) is much less efficient, as the body tries to compensate more-or-less ineffectively for rising internal temperature.
- Under normal circumstances, skin is cooler than the body core. Thus increasing or decreasing blood flow through capillaries in the papillary layer of dermis can dissipate or conserve body heat.
- Fat of the hypodermis can serve as effective insulation. Regulation of blood flow into the dermis determines the extent which this insulation is used (to conserve heat) or bypassed (to dissipate heat).
Comments and questions: dgking@siu.edu
SIUC / School of Medicine / Anatomy / David King
https://histology.siu.edu/intro/skin.htm
Last updated: 24 September 2024 / dgk